
Glycan-specific molecularly imprinted polymers towards cancer diagnostics: merits, applications, and future perspectives
Pengfei
Li
and
Zhen
Liu
 *
*
State Key Laboratory of Analytical Chemistry for Life Science, School of Chemistry and Chemical Engineering, Nanjing University, Nanjing 210023, Jiangsu, China. E-mail: zhenliu@nju.edu.cn; Fax: +86-25-8968-5639
First published on 15th January 2024
Abstract
Aberrant glycans are a hallmark of cancer states. Notably, emerging evidence has demonstrated that the diagnosis of cancers with tumour-specific glycan patterns holds great potential to address unmet medical needs, especially in improving diagnostic sensitivity and selectivity. However, despite vast glycans having been identified as potent markers, glycan-based diagnostic methods remain largely limited in clinical practice. There are several reasons that prevent them from reaching the market, and the lack of anti-glycan antibodies is one of the most challenging hurdles. With the increasing need for accelerating the translational process, numerous efforts have been made to find antibody alternatives, such as lectins, boronic acids and aptamers. However, issues concerning affinity, selectivity, stability and versatility are yet to be fully addressed. Molecularly imprinted polymers (MIPs), synthetic antibody mimics with tailored cavities for target molecules, hold the potential to revolutionize this dismal progress. MIPs can bind a wide range of glycan markers, even those without specific antibodies. This capacity effectively broadens the clinical applicability of glycan-based diagnostics. Additionally, glycoform-resolved diagnosis can also be achieved through customization of MIPs, allowing for more precise diagnostic applications. In this review, we intent to introduce the current status of glycans as potential biomarkers and critically evaluate the challenges that hinder the development of in vitro diagnostic assays, with a particular focus on glycan-specific recognition entities. Moreover, we highlight the key role of MIPs in this area and provide examples of their successful use. Finally, we conclude the review with the remaining challenges, future outlook, and emerging opportunities.
 Pengfei Li | Pengfei Li received his bachelor's degree from Tongji University in 2017. He obtained his PhD from Nanjing University under the supervision of Prof. Zhen Liu in 2023. His research is centered around the intersection of chemistry and medicine, with particular emphasis on developing innovative proteoform-resolved assays and targeted metabolomic methods for disease diagnosis and embryogenesis-associated single-cell analysis. |
 Zhen Liu | Dr Zhen Liu is a Distinguished Professor at Nanjing University. He was awarded the National Science Fund for Distinguished Young Scholars in 2014. He is a Fellow of the Royal Society of Chemistry (FRSC), an associate editor of Analytical Methods and an advisory board member of multiple journals. He was awarded a few important awards including the ACS Advances in Measurement Science Lectureship Award (2020). His major research interest is to develop biomimetic molecular recognition materials and assays for challenging applications, such as, single cell analysis, disease diagnosis, cancer therapy and viral inhibition. He has published 200+ peer-reviewed journal papers. |
1. Introduction
Cancer continues to be one of the leading cause of death worldwide,1 emphasizing the need for early detection to improve long-term survival and reduce mortality rates. Currently, extensive research is dedicated to uncovering innovative biomarkers, including proteins2,3 and nucleic acids,4,5 due to their potential in detecting tumours at an early stage, even before symptoms are noticeable. Nevertheless, the current diagnostic sensitivity and specificity of most biomarkers still do not meet satisfactory levels.6 Furthermore, numerous types of cancer still lack reliable markers for early detection.Glycans are present in every cell studied to date across the kingdoms of life.7 In mammals, all classes of biomolecules can be found in a glycosylated form, ranging from proteins,8 lipids,9 secondary metabolites10 to recently reported RNAs.11 The extensive presence of glycans in organisms emphasizes their crucial role in regulating diverse physiological and pathological functions, including cell communication, immunomodulation, embryogenesis and tumour progression.7,12 Unlike proteins and nucleic acids, the biosynthesis of glycans is not precisely defined at the genetic level.10 It is a dynamic process that depends on multiple environmental stimuli, including the local milieu of enzymes, sugar precursors and organelle structures as well as the cell types involved and cellular signals.12 Given that the development of cancer is a combination of genetic and environmental factors,13,14 in the case where disease states cannot be well reflected by genetic or proteomic markers, glycans can report on cancers much better. Virtually, the aberrations in glycan structures and glycosylation patterns have been observed in various types of cancers,15,16 and their critical role as biomarkers has been recognized by the fact that diagnosis with protein-specific glycan changes rather than only proteins will have improved clinical outcomes.17,18
Tremendous efforts have been dedicated to discovering potent glycan biomarkers. Particularly, a substantial number of aberrant glycans have been identified via advanced mass spectrometry (MS) technology.19 However, a striking discrepancy still exists between these upstream efforts and the number of diagnostic assays in clinical practice. Currently, only a few glycan markers are approved by the US Food and Drug Administration (FDA) and widely used in clinical settings, such as AFP-L3 glycoforms (core-fucosylated N-glycans that exhibit stronger affinity to lentil culinaris agglutinin) for hepatocellular carcinoma diagnosis20 and CA19-9 (a tetrasaccharide carbohydrate termed sialyl Lewis a) for pancreatic cancer diagnosis.21,22 The dismal progress in translating glycans into clinical diagnostic applications can be attributed to various reasons, including difficulties in clinical validation and regulatory considerations. However, one major technical challenge is the lack of anti-glycan antibodies with high affinity and exquisite selectivity, as glycans are poorly immunogenic and structurally similar.23,24 Although there are ongoing efforts to explore alternative recognition ligands for developing diagnostic assays, such as lectins,25 boronic acid26,27 and aptamers,28 limitations in affinity, selectivity, stability and versatility still hinder the translation of glycans in clinical diagnostics.
Molecularly imprinted polymers (MIPs), a category of artificial receptors with custom-designed binding sites for targeted molecules,29 present a viable solution to address these challenges. Specifically, customized MIPs possess the ability to selectively bind to a wide range of anomalous glycans, including N-glycans,30 O-glycans,31 glycosaminoglycans,32,33 and gangliosides,34 with remarkable affinity. This capacity holds the potential to enable clinical translation of numerous MS-identified glycans that lack specific antibodies, effectively broadening the scope and applicability of glycan-based diagnostics. Additionally, glycoform-resolved diagnostics could also be achieved through tailored cavities of MIPs,17 further improving the precision of diagnostics. Moreover, MIPs exhibited other noteworthy merits, including easy preparation, great stability, and low cost.35 These advantageous characteristics position MIPs as a promising tool for catalysing a transformative revolution in glycan-based diagnostics, ultimately facilitating long-term benefits for patients.
Although the synthetic strategies and vast biomedical applications of MIPs have been reviewed,29,35,36 a review focusing on the unique role of glycan-specific MIPs in broad and precise diagnostics is apparently lacking. In this review, we aim to introduce the promising potential of glycan-specific MIPs for developing diagnostic assays with an emphasis on their excellent binding characteristics to glycans (Fig. 1). We briefly summarize the current status of glycans as biomarkers. Next, we outline advanced glycan-specific bio- and artificial receptors and their advantages and potential drawbacks for diagnostics, without considering the substantial efforts concentrated on the chemical labelling methods to target glycans, as these methods are not suitable for clinical diagnostic assays. Then, we highlight the successful examples of glycan-specific MIPs in diagnostics, which is a very new field with less than twenty publications, making it a timely survey to introduce the topic and discuss the current challenges and future opportunities in this significant field.
 | ||
| Fig. 1 Overview of glycan-specific MIPs-based diagnostics. In comparison to traditional natural and artificial glycan binding receptors, glycan-specific MIPs exhibit several advantages in terms of generality, stability, and selectivity. These features have the potential to revolutionize glycan-based diagnostics, offering “sweeter than ever” performance. | ||
2. Cancer-associated glycan changes
Altered patterns of glycosylation are a characteristic feature of a dysfunctional cellular phenotype. This phenomenon was initially observed in the 1970s when it was shown that healthy fibroblasts have smaller membrane glycoproteins compared to their transformed counterparts.37 Subsequent studies have demonstrated that lectins exhibit varying binding affinities for healthy and malignant tissues.38 More recently, advancements in mass spectrometry have allowed for the direct characterization of disease-associated glycans,39 shedding further light on these unique molecular changes that occur during malignant transformation. Despite the discovery of numerous disease-related glycans, their validation as diagnostic markers in clinical practice remains inadequate. Therefore, in this section, we do not attempt to comprehensively summarize the vast field of aberrant glycans identified by mass spectrometry recently, but rather briefly introduce a few well-recognized glycan changes in tumours and provide some examples of tumour-associated glycan markers found in glycoproteins, glycolipids and proteoglycans, which have been validated in a significant number of clinical samples (Fig. 2). | ||
| Fig. 2 Cancer-associated glycan changes. (A) Major types of glycoconjugates in humans. (B) Main cancer-associated glycan changes found in glycoproteins, glycolipids and glycosaminoglycans. (C) FDA-approved glycan markers: AFP-L3 and CA19-9. Region with red shading represents specific glycoforms of AFP-L3. | ||
2.1 Glycoproteins
Glycoproteins comprise approximately 50% of the total cellular proteome and >90% of the secreted proteome.8 Protein glycosylation refers to the covalent attachment of carbohydrates to polypeptides, including the addition of N-linked glycans, O-linked glycans, C-linked glycans and glycosylphosphatidylinositol (GPI) anchors.40 Among these, N-linked glycans and O-linked glycans have been extensively studied due to their central roles in mediating molecular and cellular functions, ranging from protein folding to intercellular communication.Alterations in N-linked glycans typically involve an increase in core-fucosylation, sialylation and branching (Fig. 2B).41,42 On the other hand, mucin-type glycans are frequently observed in O-linked glycans associated with malignant tumours.43 Mucins belong to the class of glycoproteins that are components of mucus gel layers, playing a critical role in shaping the functions of mucus barriers.44 They can act as scaffolds for the expression of cancer-associated glycan epitopes, such as Tn antigen and sialyl Tn antigen (sTn) (Fig. 2B).45,46 Besides, certain mucins (for example, CA125 and CA153) have been widely used as diagnostic markers for cancer.47,48
In addition to aforementioned changes, overexpression of some certain terminal glycans is also commonly found. Within tumour tissue, glycosyltransferases (for example, sialyltransferases and fucosyltransferases) involved in linking terminating residues on glycans tend to be overexpressed, leading to the overexpression of certain terminal glycan epitopes, such as sialyl Lewis a (SLeA), sialyl Lewis x (SLeX), Lewis y (LeY) and polysialic acid (Fig. 2B).49–51 These glyco-neoantigens, normally associated with the extravasation of lymphocytes from the blood, can facilitate metastatic spread and serve as unique biomarkers.12,52
Overproduction of certain glycoproteins is another common feature of tumours. For example, in patients diagnosed with hepatocellular carcinoma (HCC), the concentration of alpha fetoprotein (AFP) in serum can reach extraordinarily high levels.53 While the clinical utility of serum AFP in HCC patients is widely recognized, the specificity of AFP fails to meet the needs for clinical diagnosis.54,55 This is due to the fact that elevated AFP levels can also occur in many other diseases, such as gastric cancer and testicular germ cell cancer.56,57 Moreover, approximately 30% of HCC patients show levels of AFP below the threshold (AFP < 25 ng mL−1),58 further limiting its effectiveness for clinical diagnosis. In 1981, the cancer-specific microheterogeneity of AFP was elucidated through the use of Lens culinaris agglutinin (LCA), which recognizes core fucosylated glycans from AFP.59 After that, the proportion of the LCA-reactive fraction of AFP (referred to as AFP-L3) compared to total AFP has been proposed as an improved biomarker in HCC diagnosis, particularly in differentiating HCC from cirrhosis and hepatitis.60 Nowadays, AFP-L3 has obtained approval from the FDA as a biomarker for the early detection of HCC and is widely employed in clinical practice (Fig. 2C).61
The successful application of AFP-L3 has inspired further efforts to enhance the diagnostic specificity of other cancers. Prostate cancer, being the most prevalent cancer among men in Europe and the USA,62 serves as a prime example. In 1994, the FDA approved the use of prostate-specific antigen (PSA) for detecting and diagnosing prostate cancer.63 However, the PSA-based blood test is not specific to prostate cancer, as increased PSA levels can also be caused by benign prostatic hyperplasia (BPH) or prostatitis.64,65 Moreover, patients with PSA levels between 4–10 ng mL−1 are considered to be in a diagnostic “grey zone”,62 leading to unreliable diagnostic accuracy. Therefore, in order to develop a more reliable diagnostic tool than the current PSA test, many studies have explored whether changes in PSA glycosylation patterns can differentiate between prostate cancer and BPH. Some recent reports have shown an increase of α2–3 sialic acids in PSA in men with prostate cancer compared to those with BPH.66 While further clinical validation is still needed, it is clear that glycoform-resolved diagnostic methods for PSA have potential translational value and clinical significance in the future.
Overall, tumour cells exhibit significant changes in glycoproteins, especially in N-linked and O-linked glycan structures. These changes in glycans on specific glycoproteins can serve as important diagnostic markers. Given that most of the markers approved by the FDA are glycoproteins,67 protein-specific glycan changes have enormous potential for clinical translation. However, it seems unlikely—in the near future, at least—that any single glycan change will be capable of definitively distinguishing patients from healthy ones. Therefore, diagnostic assays, or more specifically, affinity reagents, that can selectively bind multiple cancer-associated glycans will revolutionize future clinical applications significantly.
2.2 Glycolipids
As the name suggests, glycolipids are glycoconjugates of lipids and they distributed throughout animals, plants and micro-organisms. They are indispensable in maintaining the stability of the membrane. Besides, they can also act as receptors for viruses and other pathogens to enter cells.68 Glycolipids encompass a wide variety of compounds, including glycosphingolipids (cerebrosides, globosides, gangliosides and others), glycoglycerolipids, glycophosphospholipids and so on. Glycosphingolipids are one of the most abundant glycolipids in humans and are found at elevated levels in cancer tissues. For example, the increased expression of GD2, GD3, GM3 and Globo H has been reported in high-grade bladder cancer,69 small-cell lung carcinoma70 and neuroblastoma.71 In addition, tumour-associated glycan epitopes, such as SLeA, SLeX, LeY, Globo H and polysialic acid, could also be observed in glycolipids with malignancy (Fig. 2B), pinpointing their critical role as diagnostic biomarkers.The uniqueness of glycolipids as disease biomarkers stems from the wealth of structural information present in both their glycan and lipid components, which can provide insights into various disease states. However, the high structural heterogeneity of glycolipids also poses a challenge in developing a universal extraction method that can efficiently isolate all types of glycolipids. Therefore, optimizing extraction protocols is necessary to achieve accurate diagnostic applications.
2.3 Glycosaminoglycans
Glycosaminoglycans (GAGs) are a group of naturally occurring heteropolysaccharides found in all mammalian tissues. They can exist as part of proteoglycans or as independent chains. Based on the specific monosaccharides and their sulfation patterns, GAGs can be classified into several major categories, including hyaluronan (HA), chondroitin sulfate (CS) and dermatan sulfate (DS), heparin and heparan sulfate (HS), and keratan sulfate (KS).Studies have shown elevated levels of glycosaminoglycans in various types of cancer (Fig. 2B).72,73 For example, Bratulic et al. conducted a study on urine and plasma samples to analyze glycosaminoglycan profiles (GAGomes) as potential biomarkers for the early detection of 14 different types of cancer.74 2064 samples from 1260 cancer or healthy subjects were collected and cancer-specific changes in biofluidic GAGomes were observed. Their findings showcased the potential of free glycosaminoglycans as reliable and accessible biomarkers for cancer detection, regardless of the stage. A similar study by Zhang et al. focused on the use of oncofetal chondroitin sulfate (ofCS) as a plasma biomarker for pan-cancer detection.75 Through an enzyme-linked immunosorbent assay (ELISA), they optimized the ratiometric detection of ofCS glycans and its protein backbone in plasma samples. The results showed significantly higher detection levels in cancer patients, and validation studies with two independent cohorts covering 11 different malignant tumours indicated the potential of ofCS as a pan-cancer biomarker.
Despite the promising potential of glycosaminoglycans as biomarkers, there is a limitation that needs to be considered. GAGs are present in various tissues and biological fluids such as the extracellular matrix, cartilage, skin, and blood. This widespread distribution makes it challenging to determine the specific tissue origin of GAG biomarkers and their relevance to specific diseases or conditions. This also explains why they are commonly used as pan-cancer biomarkers. While pan-cancer diagnosis is useful for initial health screening, the lack of specificity may restrict its diagnostic utility.
3. Traditional glycan binding receptors
Due to the complexity of biological systems and samples, the recognition of glycans plays a critical role in the development of reliable diagnostic methods in the fields of medicine. However, this recognition task is challenging owing to the diverse and complex glycan structures, which manifest vast heterogeneity in terms of composition, linkage, branching, conformation, and modification.76 In addition, entities that bind to glycans must discriminate subtle differences among a broad array of similar structures, such as the stereochemistry of a single hydroxyl group, which is quite a high-level requirement for exquisite recognition. Moreover, the strong solvation of glycans in water represents another major hurdle. Receptors need to compete with an extensive network of hydrogen bonds that surround the glycans, adding another layer of difficulty to their specific recognition.Despite the challenges faced in glycan recognition, significant efforts are being made to advance this field. For example, Yamashina et al. have developed a coordination-driven molecular capsule that effectively captures D-sucrose in water.77 Through theoretical calculations and control experiments, they have demonstrated that the unique selectivity of this capsule is due to multiple CH–π interactions between the sucrose hydrocarbon backbone and the shape-complementary polyaromatic cavity of the capsule. This provides important insights for the design and development of glycan-binding receptors. Similarly, Xiong et al. have synthesized a silica gel modified with glucopyranoside-Schiff base, which exhibits impressive selectivity and high adsorption capacity for sialylated glycans.78 Using this artificial receptor, they have successfully identified novel sialylation sites from complex bio-samples, greatly advancing our understanding of sialylated glycan-related cellular biological events. These developments are undoubtedly exciting. However, it is important to recognize that the widespread application of these glycan-binding receptors still needs further exploration. Currently, prevalent glycan binding receptors encompass antibodies, lectins, boronic acids and aptamers. These well-established receptors will be the main focus of discussion in this section, as they have been widely studied and recognized for their significant roles in glycan recognition. Extensive reviews have shed light on their binding characteristics, notably affinity and selectivity.79 Nevertheless, it is imperative to acknowledge that certain clinical diagnostic applications necessitate a more comprehensive assessment of factors beyond these primary attributes. In accordance with criteria for clinical test development and adoption,80 several other crucial considerations must be taken into account: (1) reproducibility stands as one critical factor, as it determines the receptors' suitability for diverse applications, ensuring robustness across a broad range of scenarios; (2) stability assumes a key role in the storage and practical use of these entities, safeguarding their effectiveness over time; (3) affordability emerges as a prerequisite, broadening accessibility for researchers and industry professionals, ultimately fostering widespread adoption; (4) conjugation flexibility arises as a key determiner associated with the versatility of these receptors to couple with various detection methods, thereby facilitating multiplexed and sensitive analysis; (5) lastly, generality assumes an essential role, influencing the number of potential glycan biomarkers that can successfully transition into clinical practice. Therefore, these factors, along with affinity and selectivity, should be carefully considered in the evaluation and selection of glycan-binding receptors for specific clinical diagnostic applications (Fig. 3 and Table 1).
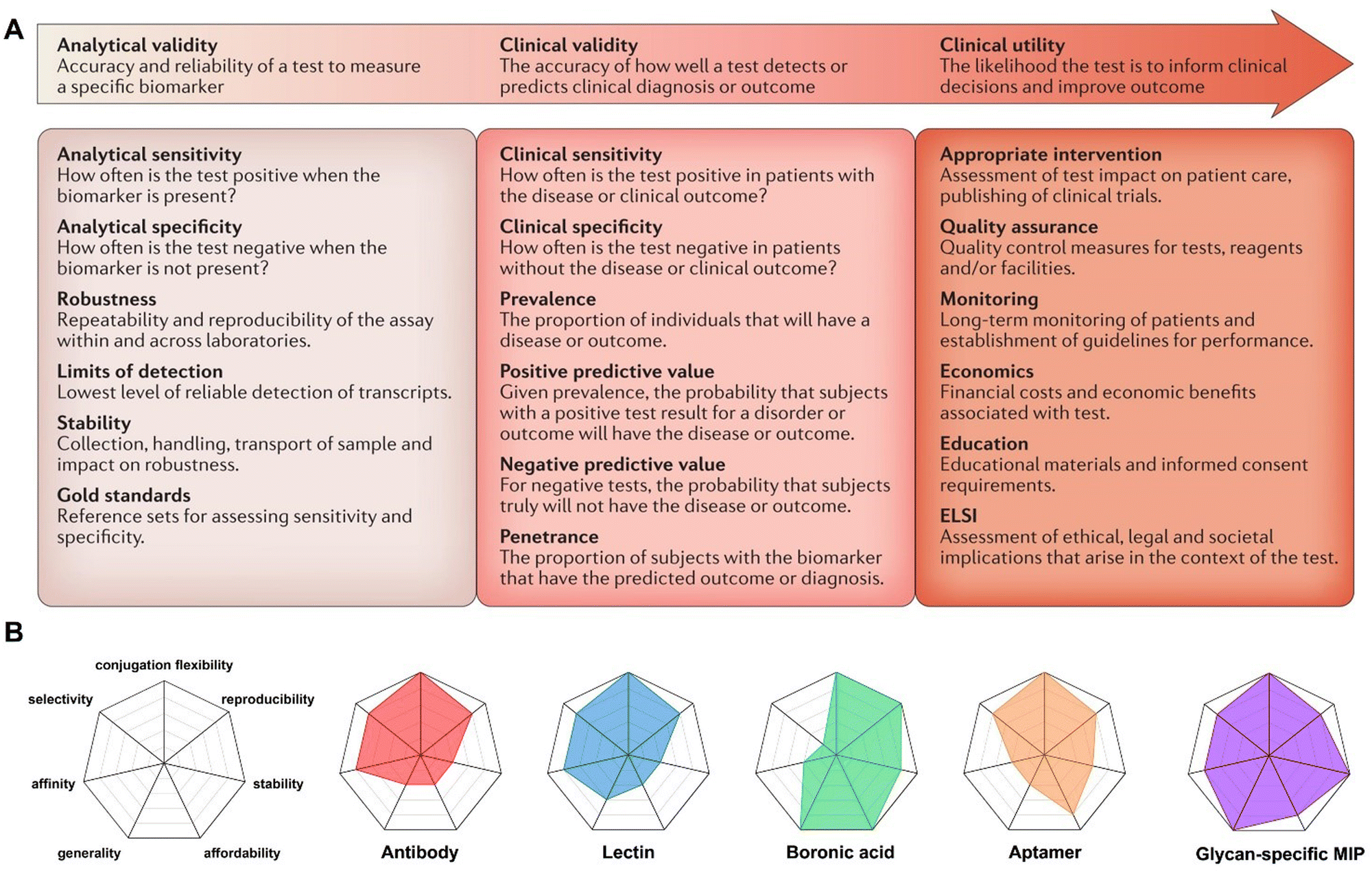 | ||
| Fig. 3 Factors that should be carefully considered in the evaluation and selection of glycan binding entities for specific clinical diagnostic applications. (A) Criteria for clinical test development and adoption. Reproduced with permission.80 Copyright 2016, Springer Nature. ELSI, National Human Genome Research Institute's Ethical, Legal and Social Implications Research Program. (B) The radar plot depicts the critical factors of different receptors. The greatest degree of desirability is located at the outermost level and is scored as 5, while the inner levels indicating the less-desired states with a score of 0. | ||
| Generality | Affinity | Selectivity | Conjugation flexibility | Reproducibility | Stability | Affordability | |
|---|---|---|---|---|---|---|---|
| Antibody | Limited anti-glycan antibody,24,79 scored 2 | Micromolar level,86,87 scored 4 | Relatively high selectivity for specific glycan structures, scored 4 | Ease of conjugation through Fc glycan remodeling,91 scored 5 | High experimental reproducibility ensured by commercialized reagent purity, scored 4 | Requirement for proper preservation of biological agents, scored 2 | High cost associated with production,84 scored 2 |
| Lectin | Ability to bind multiple targets,111–113 scored 3 | Variability with potential for optimization,100–106 scored 4 | Variability with potential for optimization,100–106 scored 4 | Ease of conjugation through chemical methods,114 scored 5 | High experimental reproducibility ensured by commercialized reagent purity, scored 4 | Requirement for proper preservation of biological agents,99 scored 2 | High costs associated with product purification and protein engineering, scored 2 |
| Boronic acid | Binding to all cis-diol containing molecules,115 scored 5 | Low affinity towards glycans at millimolar level,115 scored 2 | Binding to diverse cis-diols without discrimination,115 scored 1 | Ease of modification, scored 5 | High experimental reproducibility ensured by chemicals with analytical purity, scored 5 | Potential susceptibility to hydrolysis or degradation,116 scored 4 | Affordable cost, scored 5 |
| Aptamer | Limited glycan-specific aptamers,79 scored 2 | Potential for enhancement to micromolar level with most exhibiting low affinity,79,122 scored 2 | Relatively high selectivity for specific glycan structures, scored 4 | Ease of conjugation through chemical methods, scored 5 | High experimental reproducibility, scored 4 | Susceptibility to degradation or loss of activity over time,123 scored 3 | Cost-effectiveness when compared to biological receptors, scored 4 |
| Glycan-specific MIP | Broad generality,30–34 scored 5 | Comparability to antibodies, with potential superiority to boronic acids and aptamers,132–135 scored 4 | Glycoform-resolved specificity,17,132 scored 4 | Ease of functionalization for different purposes, scored 5 | Fine optimization of selectivity and affinity to ensure high experimental reproducibility, scored 4 | Excellent stability,131 scored 5 | Cost-effectiveness when compared to biological receptors, scored 4 |
3.1 Antibodies
Antibodies, or immunoglobulins, are specialized proteins produced by the immune system in response to foreign substances called antigens. The generation of human glycan-specific antibodies has been primarily carried out in murine systems, but with limited success. One significant obstacle is the similarity between human and murine glycomes,81 which may lead to self-tolerance and hinder the antigenicity of many human glycans in murine models. To overcome this challenge, efforts have been directed toward alternative hosts such as lampreys82 and avians,83 resulting in generating high-affinity anti-glycan antibodies. Despite progress, generating glycan-specific antibodies remains challenging due to the poor immunogenicity of glycans and the similarity among various glycan sequences. Consequently, relatively few antibodies against defined glycans are available.24 Moreover, the complex production processes contribute to the generally higher costs associated with antibody production,84 further hampering the comprehensive development of diagnostic methods.Antibodies have played a pivotal role in the discovery of specific tumour-associated carbohydrate antigens. A notable instance is the identification of CA 19-9 by Koprowski in 1979, employing a mouse monoclonal antibody in a colorectal carcinoma cell line.85 These antibodies have demonstrated exceptional selectivity for such antigens. However, it is worth noting that anti-glycan antibodies generally exhibit lower affinities, with equilibrium dissociation constant (Kd) values in the micromolar level for monovalent interactions.86,87 In contrast, protein-specific antibodies typically possess Kd values on the order of nanomolar or even picomolar.88
When employing glycan-specific antibodies, we must bear in mind that antibodies are themselves glycosylated. The glycoforms present on the Fc domains of IgG play a significant role in regulating the binding to Fcγ receptors, thus modulating the effector functions of antibodies and contributing to inflammatory pathologies (Fig. 4).12,89,90 Notably, antibodies can be easily conjugated with various labels or reporter molecules, such as fluorescent dyes, enzymes, or nanoparticles, through Fc glycan remodeling.91 This flexibility in conjugation enables antibodies to be effectively utilized alongside diverse detection methods and signal amplification strategies in diagnostic applications. However, it is important to be aware that the glycans present in the antibody may sometimes interfere with the detection process when constructing diagnostic methods. For instance, in the widely used ELISA methods, N-glycans found on the heavy chains of a capture antibody may impede the recognition of a labelling antibody towards the glycan target, thus potentially decreasing the robustness of the diagnostic methods.
 | ||
| Fig. 4 Antibodies are themselves glycosylated and the glycoforms present on antibodies contribute to inflammatory pathologies. (A) Immunoglobulin G (IgG) Fc region contains two N-glycans, one per heavy chain, attached at Asn297. These glycans play an important role in binding to Fcγ receptors. (B) IgG glycoforms with Gal-deficient glycans are pro-inflammatory. (C) IgG glycoforms with sialylated glycans are considered to be anti-inflammatory. Reprinted with permission.12 Copyright 2019, Springer Nature. | ||
In fact, glycan-specific antibodies have found extensive application in clinical practice and have been effectively validated for their binding characteristics in cancer diagnostics.92,93 Although there have been more than 1000 reported glycan-specific antibodies, a substantial portion of them target the same glycan structures, such as Tn and sTn.94,95 By some estimates, the number of glycans that could be recognized by these antibodies is only one-quarter of the total number of antibodies available.96 This redundancy significantly restricts the overall diversity of glycans that can be specifically identified and targeted. Moreover, numerous cancer-associated glycans still lack corresponding antibodies capable of effectively targeting ability.79 This deficiency in antibody coverage creates a substantial hurdle in the broader adoption of glycan biomarkers in clinical settings, subsequently impeding progress in clinical translation. Therefore, it could be foreseen that addressing this limitation in the generality of anti-glycan antibodies would be a crucial breakthrough required for their future clinical applications.
3.2 Lectins
Lectins are non-enzymatic proteins that specifically bind to carbohydrates. They are widely used in various fields, including the study of glycan patterns, the detection of glycan alterations in disease, and the exploration of structure–function relationships.97,98 Although many lectins are available for commercial use, they can be quite expensive due to the costs associated with natural product purification and recombinant protein engineering. Unlike antibodies, lectins are more resistant to heat denaturation.99 However, lectins still require careful handling, storage, and transportation to maintain their bioactivity. Any deviation from the optimal conditions, such as pH variations or exposure to denaturing agents, reducing agents, or proteases, can result in the denaturation or precipitation of lectins. Therefore, it is crucial to consider the limitations of affordability and stability when utilizing lectins in diagnostics.Lectins exhibit varying affinity and specificity towards their targets, influenced by factors such as glycan density and presentation.100–103 Generally, lectins have weak affinities for glycans and exhibit poor specificities due to the shallow binding pockets on their surfaces. To enhance their binding properties, researchers have turned to lectin engineering techniques such as site-directed mutagenesis104 and random mutagenesis.105 For instance, Peanut agglutinin is a crucial lectin in clinical settings owing to its high affinity for the tumour-associated Thomsen–Friedenreich antigen (T-antigen). To design a practical probe with improved specificity for T-antigen, Adhikari et al. employed site-directed mutagenesis by focusing on Asn41 and successfully created a mutant variant N41Q, which exhibited a four-fold increase in binding affinity for T-antigen.106 In addition to improving binding abilities, lectin engineering can also enhance their stability. Galectin-9 is a novel immune modulator comprising two carbohydrate-recognition domains and is susceptible to proteolysis due to a lengthy and flexible linker peptide between the two carbohydrate-recognition domains. Nishi et al. overcame this limitation by preparing mutant proteins through the successive truncation of the linker peptide, resulting in a highly stable mutant called G9Null that lacked the entire linker peptide while retaining its biological activities.107 However, despite the potential for engineering lectins to make them more suitable for diagnostic purposes, it should be noted that challenges remain. These include operational complexities, selection of appropriate template lectins, and construction of mutagenesis libraries. Furthermore, the lack of a widely unified method for lectin engineering poses further hindrances to their clinical utility.
Typically, a multivalent presentation of both lectin and glycans is needed to achieve physiologically relevant binding characteristics.108,109 However, in diagnostic applications where lectins are immobilized on extraction substrates, the interactions between lectins and targets are usually monovalent, resulting in significantly lower affinity and bad analytical performance. This compromised analytical performance could potentially impede the further progress in diagnostic applications involving lectins. Additionally, it is important to note that certain lectins can bind not only glycans but also non-carbohydrate ligands, such as porphyrin and peptides.110 Therefore, when using lectins for the diagnosis of glycan markers, there is a possibility of false positive results due to the presence of these non-carbohydrate ligands.
Despite the drawbacks mentioned, lectins still hold great value in clinical applications. As natural ligands, lectins have the ability to bind to multiple targets, ranging from monosaccharides111 and oligosaccharides112 to glycans.113 Furthermore, lectins can be easily conjugated to different labels using various chemical methods,114 providing numerous opportunities for further clinical utility.
3.3 Boronic acids
Boronic acids have been widely used in the covalent binding of cis-diols present in glycans. This interaction is reversible and pH-controlled. In brief, when the pH is equal to or greater than the pKa value of the boronic acid, the boronic acid can form cyclic esters with cis-diols. Conversely, when the pH is significantly lower than the pKa value of the boronic acid, the boronic acid-cis-diol complex dissociates (Fig. 5).115 | ||
| Fig. 5 Interaction between boronic acids and cis-diol-containing compounds. Reprinted with permission.115 Copyright 2017, American Chemical Society. | ||
Boronic acids are commonly preferred due to their affordable cost as compared to antibodies. Moreover, boronic acids offer the advantage of being easily modifiable, which allows for flexible detection strategies in glycan-based applications. However, their limitations are also worth noting. First, while there is a wide range of commercially available boronic acids and derivatives, only a limited subset can provide a biocompatible binding pH suitable for biological samples.115 The scarcity of boronic acids capable of operating within the physiological pH range poses a significant challenge for robust and reliable diagnostic analysis. Second, some boronic acids may be prone to hydrolysis or degradation,116 limiting their stability for clinical applications. Third, boronic acids inherently exhibit lower affinity towards glycans, which impacts their efficiency in accurately detecting and quantifying specific glycan structures. Although strategies such as multivalent binding117,118 and confinement effects119 have been explored to enhance their affinities, employing these approaches may introduce added complexity to the overall diagnostic process. Lastly, boronic acids could bind diverse cis-diol-containing compounds without discrimination. This property is advantageous for selectively enriching cis-diols from complex matrices.120 However, when it comes to reporting precise information in diagnostic methods, the lack of specificity hinders the ability to precisely identify and differentiate between specific glycan structures.
3.4 Aptamers
Aptamers are nucleic acid molecules that possess the ability to mimic antibodies through the formation of intricate 3D structures. While some aptamers occur naturally as the ligand-binding components of riboswitches, the majority are generated in vitro and can be engineered for a specific target. In comparison to antibodies, aptamers offer several advantages, including their simplicity in production, cost-effectiveness, and consistency between batches,121 making them an excellent alternative for diagnostic applications. However, glycan-specific aptamers are restricted due to the limited interactions between carbohydrates and nucleotides.79By introducing modified bases into nucleotides, it is possible to enhance the affinity of aptamers to glycans. For example, Yoshikawa et al. conducted a study where they incorporated the aromatic heterocyclic indole moiety into DNA aptamers for specifically recognize protein glycan epitopes (Fig. 6).122 Indole was used owing to two reasons. First, tryptophan, which is an indole derivative, often establishes crucial interactions within the binding pockets of anti-carbohydrate antibodies. Second, indole has been shown to favourably bind to the electron-poor C–H bonds of carbohydrates, and complementary electronic effects contribute to driving protein-carbohydrate interactions. The aptamers developed in this study exhibited a Kd in the micromolar range towards polysaccharides, underscoring the importance of employing modified bases to enhance the affinity of aptamers for glycans.
 | ||
| Fig. 6 Selection scheme for generating indole-modified aptamers. Reprinted with permission.122 Copyright 2021, Springer Nature. | ||
Although the aforementioned example provides a potential solution to enhance the affinity between aptamers and glycans, it is crucial to acknowledge that glycan recognition utilizing aptamers is still in its infancy. The challenge lies in devising a comprehensive strategy to engineer aptamers that can effectively bind to a wide array of glycans while maintaining glycoform-level resolution. Furthermore, a notable drawback of aptamers is their susceptibility to degradation or loss of activity over time,123 thereby hindering their long-term storage and potentially compromising their clinical diagnostic applications.
4. Molecularly imprinted polymers: perfect fit for glycan-based diagnostics
Molecularly imprinted polymers (MIPs) are synthetic antibody mimics that are obtained through the polymerization of functional monomers and crosslinker molecules in the presence of a template (Fig. 7).124–126 MIPs have made significant contributions to a wide range of applications, including catalysis,127 sensing,128 and separation.129 In particular, their unique characteristics make them highly advantageous for developing clinical diagnostic methods. MIPs are generally more affordable compared to antibodies due to their synthetic nature. Moreover, they are often reusable and can be synthesized in large quantities,130 which effectively reduces production costs. This cost-effectiveness makes MIPs readily accessible to both academic and industrial communities, ultimately facilitating their extensive utilization from the preclinical to the clinical stage. In addition to their economic benefits, MIPs are renowned for their robustness and durability. They exhibit remarkable structural integrity and binding activity even when exposed to diverse conditions such as temperature variations, pH fluctuations, and protease exposure.131 Excellent stability sets MIPs apart from antibodies to offer a versatile tool for a broad spectrum of diagnostic applications. | ||
| Fig. 7 Schematic diagram of the preparation process of molecularly imprinted polymers. | ||
Compared to natural receptors, MIPs offer significant advantages primarily due to their broad generality. Reported studies have demonstrated the capability of MIPs to bind a wide range of glycans, encompassing N-glycans,30 O-glycans,31 glycosaminoglycans,32,33 gangliosides34 and so on. Importantly, MIPs exhibit selectivity and affinity towards these targets that can be comparable to antibodies and lectins.132 For aberrant glycans that lack specific antibodies or lectins, MIPs hold the potential to enable their clinical translation, thereby extending the scope and applicability of glycan-based diagnostics.
Compared to artificial ligands, MIPs provide a highly versatile platform capable of incorporating a wide range of affinity ligands, including boronic acids and aptamers. For example, MIPs can be constructed using boronate-affinity glycan-oriented surface imprinting30,133 or aptamer-molecularly imprinted polymer hybrids (Fig. 8).134,135 These advanced materials exhibit significantly improved affinity and selectivity towards targets when compared to boronic acids and aptamers used individually. More importantly, by incorporating various affinity ligands, types of interactions between MIPs and glycans can be significantly enhanced to achieve glycoform-resolved recognition. This is particularly important because glycans often exhibit subtle differences that are challenging to distinguish using traditional receptors. The ability of MIPs to discriminate specific glycoforms holds great promise in improving accuracy in diagnosis, as only certain unique glycoforms are highly associated with cancers. Thus, the versatility and effectiveness of MIPs make them a superior choice over other artificial ligands in diagnostic applications.
 | ||
| Fig. 8 MIPs provide a highly versatile platform capable of incorporating a wide range of affinity ligands, including boronic acids and aptamers. (A) Scheme of boronate-affinity surface imprinting. Reprinted with permission.133 Copyright 2017, Springer Nature. (B) Scheme of aptamer-molecularly imprinted polymer hybrids. Reprinted with permission.134 Copyright 2019, American Chemical Society. | ||
Overall, the unique properties of MIPs, such as cost-effectiveness, stability, generality and versatility, position them as exceedingly valuable tools in the realm of glycan-based diagnostics. They hold great potential in enhancing the accuracy and efficiency of diagnostic tests in various healthcare and research settings, which is indispensable for advancing this promising area. We anticipate that the development of advanced glycan-specific MIPs will pave the way for broader and more precise cancer diagnosis, opening up new avenues and opportunities for improving patient outcomes.
5. Glycan-specific MIPs in cancer diagnostics
The importance of disease diagnosis cannot be overstated, as it serves as the foundation for determining appropriate treatment strategies and predicting patient outcomes. In recent years, biomarker-based diagnosis has become a widely accepted approach, allowing healthcare professionals to gain a comprehensive understanding of diseases at the molecular level. Among the various biomarkers, aberrant glycan patterns show great potential for disease diagnosis. Given that glycans are usually found on the outermost surfaces of cellular and secreted macromolecules in biofluids,12 techniques such as imaging and liquid biopsy have gained widespread utilization in glycan-based diagnostics. Currently, there has been growing attention towards integrating glycan-specific MIPs into liquid biopsy and imaging-based diagnostic approaches. This integration can provide valuable information on the presence, progression, and molecular profile of various diseases, expanding the scope of current diagnosis and enhancing healthcare outcomes.5.1 Glycan-specific MIPs in liquid biopsy-based diagnostics
Liquid biopsy, a state-of-the-art diagnostic technology, analyses biomarkers present in bodily fluids such as blood or urine and has emerged as a promising alternative to traditional tissue biopsies. Unlike tissue biopsies, which may not accurately represent the heterogeneity and dynamic changes within a tumour, liquid biopsy offers a more comprehensive view. It captures genetic, protein, or glycan alterations from various tumour sites, providing valuable insights into tumour evolution and helping tailor personalized treatment strategies. While many studies have focused on using glycoprotein-specific MIPs in liquid biopsy-based diagnostics,136,137 it is noteworthy that glycans and glycoproteins are not synonymous. Changes in glycoprotein abundance, the degree of glycosylation, and specific glycan structures all have relevance to various diseases. Therefore, detecting glycoproteins as a whole might not fully reveal the valuable information hidden in glycans. As a result, we will not discuss glycoprotein-specific MIPs in this section.Despite extensive research on the preparation of glycan-specific MIPs,30–34 their incorporation into liquid biopsy-based diagnostics has not been widely explored. This could be attributed to the challenges involved in obtaining pure glycan templates that are specifically associated with diseases and the incompatibility of traditional MIPs, which are often hydrophobic and unsuitable for aqueous-based biological applications.
Recently, with advancements in glycobiology and molecular imprinting technology, an increasing number of glycan-specific MIPs are being utilized in diagnostics (Table 2). One common target in this field is carbohydrate antigens, such as CA125,138,139 CA19-9,140 and CA153.141 For instance, Li et al. investigated the use of glycan epitope sialyl Lewis A as a template for the development of MIP sensors for the diagnosis of pancreatic adenocarcinomas (Fig. 9A).140 The resulting MIP sensors exhibited an excellent linear response to CA19-9 within the range of 0.1–5 U mL−1, with a detection limit of 0.028 U mL−1. Furthermore, the analysis of serum samples showed a strong correlation with results obtained from an established ELISA method, highlighting the promising potential of glycan-specific MIPs in diagnostics. However, certain terminal glycan epitopes can also be expressed on various other disease-associated proteins, limiting the diagnostic specificity when using only glycan-specific MIPs to capture biomarkers.
| Marker | Characteristics of glycan-specific MIPs | Characteristics of constructed assays | Ref. | ||
|---|---|---|---|---|---|
| Maximum cross-reactivity (%) | Affinity (U mL−1) | limit of detection (U mL−1) | linear response range (U mL−1) | ||
| CA125 | 4.8 | 0.698 | 0.01 | 0.1–10 | 138 |
| CA125 | 0.775 | 0.01 | 0.01–500 | 139 | |
| CA19-9 | ca. 15 | 0.028 | 0.1–5 | 140 | |
| CA153 | 0.141 | <0.1 | 0.1–100 | 141 | |
| CEA | 15.2 | 0.0002 | 0.0169–169 | 142 | |
| CA72-4 and CA19-9 | 0.0041 and 0.0032 | 0.005–200.0 | 143 | ||
| AFP-L3 | 16.1 | 0.121–9.68 | 144 | ||
| AFP-Gm | 0.605–9.68 | 17 | |||
 | ||
| Fig. 9 Glycan-specific MIPs in liquid biopsy-based diagnostics. (A) Schematic representation of using glycan epitope sialyl Lewis A as templates for the development of MIP sensors to diagnose pancreatic adenocarcinoma. Reproduced with permission.140 Copyright 2018, Elsevier. (B) Scheme showing the simultaneous detection of CA72-4 and CA19-9 for diagnosing gastric cancer. Reproduced with permission.143 Copyright 2018, Elsevier. (C) Scheme of triple MIP-based immune-sandwich assay to detect AFP-L3/AFP for diagnosing liver cancer. Reproduced with permission.144 Copyright 2022, Royal Society of Chemistry. (D) Schem of dual-modal ratiometric immunoassay using MIPs for precise HCC diagnosis. Reproduced with permission.17 Copyright 2022, John Wiley & Sons. | ||
To address this challenge, an orthogonal dual MIP-based sandwich assay has been developed.142 In this approach, a peptide epitope-imprinted substrate is employed to capture carcinoembryonic antigen (CEA), a widely used marker for colon cancer, from serum. Simultaneously, glycan-imprinted nanoparticles are utilized as labelling nanotags to recognize glycans. This orthogonal double recognition strategy significantly improves the analytical specificity, reducing the maximum cross-reactivity from 14.4% and 15.2% for epitope recognition and glycan recognition, respectively, to 8.2% for double recognition. Moreover, by measuring the CEA level in human serum, this method effectively differentiates between colon cancer patients and healthy individuals.
Although the association between glycans and diseases has gained recognition, there remains a question about whether specific glycans hold enough information to accurately report on diseases. To enhance diagnostic accuracy, a promising approach is the transition from single to multiplex glycan biomarkers. In this regard, Luo and colleagues developed a glycan-imprinted sensor for the simultaneous detection of CA72-4 and CA19-9 (Fig. 9B).143 The sensor was created by electro-polymerization, using the distinctive sTn glycan on the surface of CA72-4 and the characteristic SLeA glycan on the surface of CA19-9 as template molecules. Additionally, two specific lectins were used to label different glycosyl chains to form a sandwich assay. The generated assay displayed high sensitivity to CA72-4 and CA19-9 in the concentration range of 0.005–200.0 U mL−1, offering a simple, rapid, and cost-effective alternative for accurately diagnosing gastric cancer. However, it is important to note that simultaneously imprinting two types of glycans can increase the heterogeneity of the imprinted cavity, thus impacting the analytical specificity. While using two glycan targets could improve diagnostic specificity, these conflicting factors can influence the ultimate performance.
Another promising approach to enhance diagnostic accuracy is the use of the proportion of disease-associated glycans to total glycoproteins as an improved biomarker. As we mentioned previously, AFP-L3/AFP has been widely utilized in diagnosing HCC. To replace antibodies and lectins with MIPs, Pang et al. developed a triple MIP-based immunosandwich assay (triMIP-PISA) to detect AFP-L3/AFP (Fig. 9C).144 An N-terminal peptide-imprinted MIP was employed to capture AFP from serum, while a C-terminal peptide-imprinted MIP and a fucose-imprinted MIP were used to specifically recognize the AFP and AFP-L3 fraction. By facilitating simultaneous plasmon-enhanced Raman detection of AFP and AFP-L3 in serum, HCC patients could be accurately distinguished from healthy individuals. The triMIP-PISA method demonstrated enhanced precision compared to an antibody plus lectin-based immunofluorescence assay.
According to the examples reported above, glycan-specific MIPs have shown promising results in detecting biomarkers such as CA19-9 and AFP-L3. However, it should be emphasized that antibodies or lectins are already available for these biomarkers. The key advantage of glycan-specific MIPs should go beyond competing with antibodies or lectins for similar or improved results. Therefore, the attractive direction in designing relevant diagnostic methods should focus on translating potential glycan biomarkers without antibodies or lectins into clinical practice. To this end, Li et al. developed a dual-modal ratiometric immunoassay using MIPs for precise HCC diagnosis (Fig. 9D).17 They identified a set of glycans, referred to as Gm, which consist of a series of core-fucosylated and/or sialylated N-glycans and have a stronger association with HCC compared to AFP-L3. By constructing a “pit one against ten” strategy to develop Gm-specific MIP, a new glycoform-resolved immunosandwich assay was formed and AFP-Gm/AFP was detected. The comparison between AFP-Gm/AFP and AFP-L3/AFP demonstrated improved precision, highlighting the significant value of glycan-specific MIPs in developing glycoform-resolved diagnostic methods for potential clinical translation.
Glycan-specific MIPs have shown significant potential as artificial receptors in diagnostic assays. Integrating MIPs into liquid biopsy-based diagnostics offers several advantages, encompassing increased precision and cost-effectiveness. However, there is still room for further improvement in this field. One area that requires attention is the existing processing methods, which can be complex and time-consuming. To make these diagnostic assays more practical for widespread use, high-throughput and mechanized operations need to be developed. This would enable faster and more efficient acquisition of diagnostic results. Furthermore, investing additional effort in controllable MIP preparation, cross-reactivity verification, and other related processes is crucial. These steps are necessary to ensure robust and reliable diagnostic outcomes. By addressing these aspects, the diagnostic assays utilizing MIPs can become more reliable and effective in clinical settings. It is also worth noting that the potential of MIPs has not been fully explored yet. To advance this field further, more research should be focused on the use of glycan-specific MIPs to target glycans without antibodies or lectins. Additionally, more advanced MIPs that could enable glycoform-resolved recognition should be developed to enhance the sensitivity and specificity of glycan-based diagnostic methods. These advancements can facilitate the translation of glycan-based diagnostics into routine clinical practice.
5.2 Glycan-specific MIPs in imaging-based diagnostics
Liquid biopsy has shown promise in the analysis of bodily fluids, offering a minimal-invasive method for cancer diagnosis. However, there are certain types of cancer for which no bodily fluid markers are currently available, such as lung cancer. In these cases, it becomes essential to directly analyse cells to obtain accurate diagnostic information. Consequently, cellular imaging-based diagnostics have been gaining increasing attention in recent years, because of their ability in translating biological information into visual signals, providing valuable insights into the bio-distributions, dynamic changes, and expression levels of abnormal tumour-specific glycans. Existing imaging-based diagnostic methods rely on cellular analysis using lectins or monoclonal antibodies.145,146 However, these approaches have limitations in terms of sensitivity, selectivity and cost, hindering their wider application. A fascinating and promising future direction in this field is to target aberrant glycosylation using glycan-specific MIPs. This approach holds great potential for improving the effectiveness and accuracy of cancer diagnosis.Recently, imaging-based diagnostics have primarily focused on targeting specific monosaccharides on the surface of cells, such as sialic acid147–150 and glucuronic acid.151,152 Sialic acid, positioned at the end of glycans, plays a critical role in regulating innate immunity. Extensive research has established a clear connection between higher sialic acid levels and metastatic cancer cells.153 To further investigate the expression levels of sialic acid on cancer cells, a recent study employed MIPs tailored specifically for sialic acid for imaging four distinct chronic lymphocytic leukemia (CLL) cell lines.147 Interestingly, the more aggressive CLL cells, HG3 and CI, exhibited significantly higher levels of sialic acid when compared to the other cell lines, Wa-osel and AIII, suggesting that sialic acid-specific MIPs have potential in screening and identifying circulating tumour cells across various stages. Moving beyond sialic acid, glucuronic acid, a vital component of hyaluronan, has garnered attention due to its association with tumour progression in humans. Hyaluronan is the most abundant glycosaminoglycan found in mammalian tissue. To explore the role of glucuronic acid, Demir et al. developed carbon dot-coated molecularly imprinted polymers (CD-MIPs) using glucuronic acid as a template (Fig. 10A).151 These CD-MIPs were used to target and visualize hyaluronan in human cervix adenocarcinoma cells (HeLa) and human keratinocytes (HaCaT). The results revealed that the HeLa cells exhibited approximately four times more bound CD-MIPs compared to the HaCaT cells, indicating that CD-MIPs are capable of distinguishing between healthy cells and tumour cells.
 | ||
| Fig. 10 Glycan-specific MIPs in imaging-based diagnostics. (A) Scheme of development of carbon dot-coated molecularly imprinted polymers with hyaluronan as a template. These MIPs were used to target and visualize hyaluronan in human cervix adenocarcinoma cells and human keratinocytes. Reproduced with permission.151 Copyright 2018, American Chemical Society. (B) Scheme showing monosaccharide-specific MIPs-based multiplex imaging to target not only sialic acid but also fucose and mannose. Pattern recognition algorithms have been employed to identify similarities and differences among various cell lines, allowing for the recognition of not only cancer cells from normal cells but also different types of cancer cells. Reproduced with permission.154 Copyright 2017, American Chemical Society. (C) An example of Gd-doped MIPs for targeting Tn antigens on cellular surface. Reproduced with permission.156 Copyright 2021, Elsevier. | ||
Despite the progress made in monosaccharide-specific MIP-based imaging, there are still limitations in terms of sensitivity and specificity in diagnostics. To address these limitations, researchers have implemented monosaccharide-specific MIP-based multiplex imaging to target not only sialic acid but also fucose and mannose (Fig. 10B).154,155 This approach enables a more comprehensive assessment of relative expression levels of monosaccharides in cells. Additionally, pattern recognition algorithms have been employed to identify similarities and differences among various cell lines, allowing for the recognition of not only cancer cells from normal cells but also different types of cancer cells. However, despite these advancements, monosaccharide-specific MIP-based diagnostics still face challenges in meeting clinical needs. One of the major difficulties lies in identifying the specific glycan biomarkers associated with these terminal monosaccharides. The limited understanding of the molecular basis behind these biomarkers hinders the establishment of clear diagnostic criteria. Consequently, it becomes challenging to accurately interpret the imaging results and establish robust diagnostic methods.
Apart from monosaccharides, cancer-associated glycan epitopes are also commonly utilized as MIP targets in imaging-based diagnostics. Examples of such epitopes include the Tn antigen156 and the sialyl Tn antigen (sTn).157 For instance, in a study by Ren et al.,156 MIPs were prepared using glycan-oriented imprinting technology, with Gal-NAc, the glycosyl of Tn antigens, being used as a template (Fig. 10C). To enhance their imaging capabilities, Gd-doped fluorescent silicon nanoparticles were incorporated into the core of the synthesized MIPs. As a result, the MIPs demonstrated the ability for fluorescence imaging and magnetic resonance imaging. Cellular imaging experiments revealed stronger signals in cancer cells MCF-7, which express higher levels of Tn antigens, while weaker signals were observed in Tn antigens low-expressing cells MDA-MB-231 and normal cells NCTC. These findings indicate that the Tn-specific MIPs exhibit excellent performance in dual-mode imaging of cancer cells. This underscores their potential for utilization in future diagnostic applications in the field of cancer detection.
According to the examples mentioned above, the use of glycan-specific MIPs shows great promise in mapping aberrant glycans on cellular surfaces. This opens up possibilities for the development of imaging-based diagnostics. However, there are some limitations that need to be addressed for clinical translation. First, the implementation of imaging-based methods requires a high demand for professionals and specialized instruments. This may pose challenges in terms of accessibility and cost-effectiveness. Second, the diagnostic sensitivity and specificity are still limited. To ensure accurate and reliable results, a clear diagnostic criterion needs to be established and validated in large clinical samples. Additionally, while cellular imaging has shown great potential, the development of glycan-specific MIPs for tissue staining-based diagnostics has been slower than expected. More research and development efforts should be dedicated to this area. Furthermore, as discussed above, glycan-specific MIPs offer critical advantages in targeting glycans without antibodies and lectins, as well as their great performance in glycoform-resolved recognition. However, there have been no recent examples of such use in imaging-based diagnostics. Therefore, more efforts should be focused on exploring and showcasing the possibilities of this approach. Future studies should dedicate their focus to developing more advanced MIPs. These may include smaller-sized MIPs to increase imaging resolution, as well as more versatile functions to enable multiplex imaging. Additionally, integrating advanced algorithms to interpret imaging results and establish diagnostic criteria would be beneficial for further advancements in this field.
6. Challenges and perspectives
6.1 Grand challenges
Glycan-specific MIPs have demonstrated great promise in enabling precise and broad diagnostics. The development of glycan-specific MIP-based diagnostics lies within a new and interdisciplinary field that combines materials chemistry, analytical science, biochemistry, and biomedicine. However, there are several challenging areas that need to be addressed, ranging from the controllable preparation of materials to the effective clinical application of these diagnostics. In the following sections, we will discuss these challenges and advances which have been achieved to address them.To achieve glycoform-resolved diagnosis, careful selection of functional monomers and crosslinking agents is necessary for constructing MIPs that can effectively bind to the target glycans. Drawing inspiration from lectin-glycan interactions, factors such as hydrogen bonds, van der Waals forces, electrostatic interactions and hydrophobic interactions need to be considered when designing MIPs.100,161–163 However, including more diverse interactions in MIPs also increases the risk of nonspecific adsorption of biocomponents other than the target glycans. Consequently, while the binding ability to the target may increase, a decrease in specificity often occurs. Therefore, finding a way to functionalize glycan-specific MIPs to simultaneously enhance specificity towards specific glycoforms and reduce nonspecific adsorption is crucial for their application in real samples.
While MIPs have been utilized in academic laboratories, the establishment of standardized production processes and systematic quality control standards for large-scale manufacturing remains unexplored. This exploration is vital to ensure that the reaction conditions and optimization parameters can be faithfully reproduced without compromising the performance of glycan-specific MIPs. Moreover, for commercial production, scalability considerations should extend beyond the MIP material itself and encompass assay design and fabrication methods. The integration of MIPs with advanced diagnostic assays, such as fully automated systems, presents a significant challenge that necessitates collaborative efforts from multiple disciplines including biochemistry, molecular biology, informatics, engineering, and commercial operations.
6.2 Future directions
Despite the grand challenges in the above section, glycan-specific MIPs reveal their merits and advantages in the clinical translation of diagnostics. Future development in this area will not only address existing challenges but also demonstrate their potential in compiling multidisciplinary technologies.To establish a comprehensive understanding of “dark matter” glycosylation across diverse tissues, cell types, and disease states, versatile and easily-prepared glycan-specific MIPs are essential tools for potential biomarker discovery. Beyond this, the development of glycan-specific MIPs presents a promising avenue for implementing novel diagnostics utilizing these “dark matter” glycan biomarkers. Future research should concentrate on the combination of glycan-specific MIPs with these “dark matter” glycan biomarkers to explore new diagnostic opportunities. Furthermore, while this review primarily focuses on the association between cancer and glycosylation, it is important to note that glycosylation can be implicated in various other diseases as well. Exploring the utility of these glycans in diagnosing other diseases with glycan-specific MIPs has immense clinical translational value.
To achieve the necessary sensitivity, various amplification strategies have been explored for engineering MIPs. One common approach is doping different reporters into the core of MIPs, including fluorescent dyes,156 Raman tags144 and plasmonic nanoparticles.17 The introduction of different reporters allows for integration with different detection methods and often significantly enhances sensitivity. However, each doping strategy has its advantages and limitations. For example, doping nanomaterials for ICP-MS analysis is straightforward but provides limited signal amplification. Conversely, doping Raman tags generally offer substantial signal amplification but may face interference from spontaneous Raman signals emitted by the sample, resulting in quantification errors. Therefore, it is crucial to develop multimodal MIP-based diagnostics that can meet various requirements in real-world applications while also providing self-verification capabilities. Furthermore, when coupling signal amplification strategies with MIPs, it is essential to ensure a consistent proportion of reporters doped onto each MIP. This necessitates advancements in encapsulation technology for engineering MIPs and the development of new strategies to achieve ultra-sensitive detection in the future.
POCT refers to a form of testing that is designed to be used near the patient's location. The World Health Organization has specified several key requirements for POCT, including affordability, sensitivity, specificity, user-friendliness, rapidity, robustness, equipment-free operation, and accessibility to end users.170 Glycan-specific MIPs fulfil many of these requirements, and their integration with other technologies like micro-nano fluidics or lateral flow immunoassays can further enhance the development of POCT devices. However, integrating such technologies also presents challenges, such as ensuring a bubble-free and proper flow path on the surface of MIPs, which consist of multiple nanosized cavities. Therefore, the development of MIP-based POCT devices requires intricate designs and fabrication protocols to address these challenges in future endeavours.
Recently, studies have demonstrated the role of glycosylation in the heterogeneity of high-grade serous ovarian carcinoma (HGSC).172 Mass spectrometry-based glycoproteomic analysis of 119 HGSC tissues revealed three major tumour clusters and five groups of intact glycopeptides. Additionally, a strong correlation between N-glycan structures and tumour molecular subtypes, such as the association of fucosylation with the mesenchymal subtype, was observed. Therefore, by developing MIPs specific to glycans associated with different cancer subtypes, diagnostic assays for cancer subtype classification can be constructed, yielding significant clinical impact. Furthermore, glycans are synthesized through a sequential process with the help of glycosylation enzymes. Designing MIP-based diagnostic assays that target changes in glycans may assist in cancer staging. Integration of this information with tumour size, location, and the involvement of nearby lymph nodes or distant organs can aid in selecting optimal treatment approaches.
Despite these promising directions, there are still several limitations that need to be considered. First, glycan levels can vary significantly among individuals, making it challenging to establish consistent cut-off values for different stages or types of cancer. Second, certain glycans may be elevated in several types of cancer or non-cancerous conditions, leading to false results and potentially misclassifying the types of cancers. Furthermore, cancer treatment itself can alter the glycan profile, leading to difficulties in accurately assessing the stage or progression of the disease during or after treatment. These challenges highlight the need for precision recognition of MIPs to specific glycoforms. Therefore, continued research and advancements in the specificity of glycan-specific MIPs are crucial for the successful implementation of glycan-based diagnostics in cancer subtyping and staging.
7. Conclusions
In summary, the rapid development of glycan-specific MIPs renders them applicable for the analysis of a large variety of glycan biomarkers directly in complex media with glycoform-level resolution, which promises a significant technology for diagnostics. By introducing glycan-specific MIPs into diagnostic applications, various options are opened up for precise and broad clinical translation, particularly in liquid biopsy-based and imaging-based diagnostics. Liquid biopsy-based diagnostics are useful for non-invasive profiling and monitoring of molecular alterations, while cell imaging provides detailed information about cellular and tissue-level glycan expression and localization. Depending on the specific clinical scenario and research needs, either one or both approaches may be employed to gain a comprehensive understanding of diseases (Fig. 11). | ||
| Fig. 11 A summary of this review. | ||
Although there are challenges to be overcome in terms of template acquisition, precise recognition, and large-scale preparation, glycan-specific MIP-based diagnostics still hold the potential to revolutionize the clinical translation of glycan biomarkers. The future direction for the development of glycan-specific MIP-based diagnostics lies in the diagnosis of “dark matter” glycan biomarkers, multimodal and ultrasensitive detection, multiplexing diagnosis, construction of POCT devices, cancer subtyping and staging, combination with artificial intelligence, and the integration of diagnosis and treatment. With these advancements, we anticipate that glycan-specific MIP-based diagnostics will increasingly emerge in clinical settings at an unprecedented pace and significantly offer a “sweeter than ever” revolution in glycan-based diagnostics.
Conflicts of interest
There are no conflicts to declare.Acknowledgements
We acknowledge financial support of the Key Grant (No. 21834003) from the National Natural Science Foundation of China and the Fundamental Research Funds for the Central Universities (No. 202200325).Notes and references
- R. C. Fitzgerald, A. C. Antoniou, L. Fruk and N. Rosenfeld, Nat. Med., 2022, 28, 666–677 CrossRef CAS PubMed
.
- L. M. Seijo, N. Peled, D. Ajona, M. Boeri, J. K. Field, G. Sozzi, R. Pio, J. J. Zulueta, A. Spira, P. P. Massion, P. J. Mazzone and L. M. Montuenga, J. Thorac. Oncol., 2019, 14, 343–357 CrossRef CAS
- L. Wu and X. Qu, Chem. Soc. Rev., 2015, 44, 2963–2997 RSC
- X. Han, J. Wang and Y. Sun, Genomics, Proteomics Bioinf., 2017, 15, 59–72 CrossRef CAS PubMed
- B. Rahat, T. Ali, D. Sapehia, A. Mahajan and J. Kaur, Front. Genet., 2020, 11, 844 CrossRef CAS
- G. A. Kwong, S. Ghosh, L. Gamboa, C. Patriotis, S. Srivastava and S. N. Bhatia, Nat. Rev. Cancer, 2021, 21, 655–668 CrossRef CAS
-
Essentials of Glycobiology, ed. A. Varki, et al., Cold Spring Harbor Laboratory Press, 3rd edn, 2015 Search PubMed
- R. Apweiler, H. Hermjakob and N. Sharon, Biochim. Biophys. Acta, 1999, 1473, 4–8 CrossRef CAS PubMed
- Q. Li, Y. Xie, M. Wong, M. Barboza and C. B. Lebrilla, Nat. Protoc., 2020, 15, 2668–2704 CrossRef CAS
- N. J. Agard and C. R. Bertozzi, Acc. Chem. Res., 2009, 42, 788–797 CrossRef CAS
- R. A. Flynn, K. Pedram, S. A. Malaker, P. J. Batista, B. A. H. Smith, A. G. Johnson, B. M. George, K. Majzoub, P. W. Villalta, J. E. Carette and C. R. Bertozzi, Cell, 2021, 184, 3109–3124 CrossRef CAS PubMed
- C. Reily, T. J. Stewart, M. B. Renfrow and J. Novak, Nat. Rev. Nephrol., 2019, 15, 346–366 CrossRef PubMed
- S. J. Virolainen, A. VonHandorf, K. C. M. F. Viel, M. T. Weirauch and L. C. Kottyan, Genes Immun., 2023, 24, 1–11 Search PubMed
- A. Mbemi, S. Khanna, S. Njiki, C. G. Yedjou and P. B. Tchounwou, Int. J. Environ. Res. Public Health, 2020, 17, 8089 CrossRef CAS
- R. Bangarh, C. Khatana, S. Kaur, A. Sharma, A. Kaushal, S. S. Siwal, H. S. Tuli, K. Dhama, V. K. Thakur, R. V. Saini and A. K. Saini, Biotechnol. Adv., 2023, 66, 108149 CrossRef CAS
- K. T. Schjoldager, Y. Narimatsu, H. J. Joshi and H. Clausen, Nat. Rev. Mol. Cell Biol., 2020, 21, 729–749 CrossRef CAS PubMed
- P. Li, J. Pang, S. Xu, H. He, Y. Ma and Z. Liu, Angew. Chem., Int. Ed., 2022, 61, e202113528 CrossRef CAS
- Y. Haga and K. Ueda, Glycoconjugate J., 2022, 39, 303–313 CrossRef CAS
- O. Aizpurua-Olaizola, J. Sastre Toraño, J. M. Falcon-Perez, C. Williams, N. Reichardt and G. J. Boons, TrAC, Trends Anal. Chem., 2018, 100, 7–14 CrossRef CAS
- H. H. S. Food and Drug Administration, Fed. Regist., 2005, 70, 57748–57750 Search PubMed
- U. K. Ballehaninna and R. S. Chamberlain, J. Gastrointest. Oncol., 2011, 3, 105–119 Search PubMed
- J. Yang, R. Xu, C. Wang, J. Qiu, B. Ren and L. You, Cancer Commun., 2021, 41, 1257–1274 CrossRef PubMed
- K. A. Janes, Sci. Signaling, 2020, 13, eaaz8130 CrossRef CAS
- Z. Polonskaya, P. B. Savage, M. G. Finn and L. Teyton, Curr. Opin. Immunol., 2019, 59, 65–71 CrossRef CAS
- P. Syed, K. Gidwani, H. Kekki, J. Leivo, K. Pettersson and U. Lamminmäki, Proteomics, 2016, 16, 1257–1265 CrossRef CAS PubMed
- D. Li, Y. Chen and Z. Liu, Chem. Soc. Rev., 2015, 44, 8097–8123 RSC
- X. Wu, Z. Li, X.-X. Chen, J. S. Fossey, T. D. James and Y.-B. Jiang, Chem. Soc. Rev., 2013, 42, 8032–8048 RSC
- W. Sun, L. Du and M. Li, Curr. Pharm. Des., 2010, 16, 2269–2278 CrossRef CAS
- K. Haupt, P. X. Medina Rangel and B. T. S. Bui, Chem. Rev., 2020, 120, 9554–9582 CrossRef CAS
- Z. Bie, Y. Chen, J. Ye, S. Wang and Z. Liu, Angew. Chem., Int. Ed., 2015, 54, 10211–10215 CrossRef CAS PubMed
- Z. Bie, R. Xing, X. He, Y. Ma, Y. Chen and Z. Liu, Anal. Chem., 2018, 90, 9845–9852 CrossRef CAS
-
M. Panagiotopoulou, S. Kunath, K. Haupt and B. Tse Sum Bui, in Synthetic Antibodies: Methods and Protocols, ed. T. Tiller, Springer New York, New York, NY, 2017, pp. 399–415 Search PubMed
- Y. Yoshimi, D. Oino, H. Ohira, H. Muguruma, E. Moczko and S. A. Piletsky, Sensors, 2019, 19, 2415 CrossRef CAS PubMed
- Z. Guo, M. Zhao and Z. Liu, Microchim. Acta, 2022, 189, 289 CrossRef CAS
- J. Xu, H. Miao, J. Wang and G. Pan, Small, 2020, 16, e1906644 CrossRef PubMed
- Y. Zhao, J. Mater. Chem. B, 2022, 10, 6607–6617 RSC
- E. Meezan, H. C. Wu, P. H. Black and P. W. Robbins, Biochemistry, 1969, 8, 2518–2524 CrossRef CAS PubMed
- G. A. Turner, Clin. Chim. Acta, 1992, 208, 149–171 CrossRef CAS
- Z. Shriver, S. Raguram and R. Sasisekharan, Nat. Rev. Drug Discovery, 2004, 3, 863–873 CrossRef CAS PubMed
- M. M. Fuster and J. D. Esko, Nat. Rev. Cancer, 2005, 5, 526–542 CrossRef CAS PubMed
- J. W. Dennis, S. Laferté, C. Waghorne, M. L. Breitman and R. S. Kerbel, Science, 1987, 236, 582–585 CrossRef CAS
- Y. J. Kim and A. Varki, Glycoconjugate J., 1997, 14, 569–576 CrossRef CAS
- M. A. Hollingsworth and B. J. Swanson, Nat. Rev. Cancer, 2004, 4, 45–60 CrossRef CAS
- S. A. Malaker, N. M. Riley, D. J. Shon, K. Pedram, V. Krishnan, O. Dorigo and C. R. Bertozzi, Nat. Commun., 2022, 13, 3542 CrossRef CAS PubMed
- M. Sletmoen, T. A. Gerken, B. T. Stokke, J. Burchell and C. F. Brewer, Glycobiology, 2018, 28, 437–442 CrossRef CAS PubMed
- R. Beatson, G. Maurstad, G. Picco, A. Arulappu, J. Coleman, H. H. Wandell, H. Clausen, U. Mandel, J. Taylor-Papadimitriou, M. Sletmoen and J. M. Burchell, PLoS One, 2015, 10, e0125994 CrossRef PubMed
- M. J. Duffy, D. Evoy and E. W. McDermott, Clin. Chim. Acta, 2010, 411, 1869–1874 CrossRef CAS
- C. Chen, Q. Chen, Q. Zhao, M. Liu and J. Guo, Ann. Clin. Lab. Sci., 2017, 47, 260–263 CAS
- S.-I. Hakomori and Y. Zhang, Chem. Biol., 1997, 4, 97–104 CrossRef CAS PubMed
- J. Taylor-Papadimitriou and A. A. Epenetos, Trends Biotechnol., 1994, 12, 227–233 CrossRef CAS PubMed
- T. F. Orntoft and E. M. Vestergaard, Electrophoresis, 1999, 20, 362–371 CrossRef CAS
- L. Oliveira-Ferrer, K. Legler and K. Milde-Langosch, Semin. Cancer Biol., 2017, 44, 141–152 CrossRef CAS
- J. D. Yang, P. Hainaut, G. J. Gores, A. Amadou, A. Plymoth and L. R. Roberts, Nat. Rev. Gastroenterol. Hepatol., 2019, 16, 589–604 CrossRef PubMed
- I. Ikai, S. Arii, M. Okazaki, K. Okita, M. Omata, M. Kojiro, K. Takayasu, Y. Nakanuma, M. Makuuchi, Y. Matsuyama, M. Monden and M. Kudo, Hepatol. Res., 2007, 37, 676–691 CrossRef
- S. Imura, H. Teraoku, M. Yoshikawa, D. Ishikawa, S. Yamada, Y. Saito, S. Iwahashi, T. Ikemoto, Y. Morine and M. Shimada, Int. J. Clin. Oncol., 2018, 23, 98–103 CrossRef PubMed
- H. Shimada, T. Noie, M. Ohashi, K. Oba and Y. Takahashi, Gastric Cancer, 2014, 17, 26–33 CrossRef CAS PubMed
- M. J. Murray, R. A. Huddart and N. Coleman, Nat. Rev. Urol., 2016, 13, 715–725 CrossRef CAS
- N. Tsuchiya, Y. Sawada, I. Endo, K. Saito, Y. Uemura and T. Nakatsura, World J. Gastroenterol., 2015, 21, 10573–10583 CrossRef CAS PubMed
- J. Breborowicz, A. Mackiewicz and D. Breborowicz, Scand. J. Immunol., 1981, 14, 15–20 CrossRef CAS
- E. Miyoshi, K. Moriwaki, N. Terao, C.-C. Tan, M. Terao, T. Nakagawa, H. Matsumoto, S. Shinzaki and Y. Kamada, Biomolecules, 2012, 2, 34–45 CrossRef CAS
- D. Li, T. Mallory and S. Satomura, Clin. Chim. Acta, 2001, 313, 15–19 CrossRef CAS
- S. Gilgunn, P. J. Conroy, R. Saldova, P. M. Rudd and R. J. O'Kennedy, Nat. Rev. Urol., 2013, 10, 99–107 CrossRef CAS
- S. D. Mikolajczyk, Y. Song, J. R. Wong, R. S. Matson and H. G. Rittenhouse, Clin. Biochem., 2004, 37, 519–528 CrossRef CAS PubMed
- B. Chughtai, J. C. Forde, D. D. M. Thomas, L. Laor, T. Hossack, H. H. Woo, A. E. Te and S. A. Kaplan, Nat. Rev. Dis. Primers, 2016, 2, 16031 CrossRef
- S. Azab, A. Osama and M. Rafaat, Transl. Androl. Urol., 2012, 1, 148–152 Search PubMed
- R. Saldova, Y. Fan, J. M. Fitzpatrick, R. W. G. Watson and P. M. Rudd, Glycobiology, 2010, 21, 195–205 CrossRef PubMed
- M. J. Kailemia, D. Park and C. B. Lebrilla, Anal. Bioanal. Chem., 2017, 409, 395–410 CrossRef CAS PubMed
- L. Nguyen, K. A. McCord, D. T. Bui, K. M. Bouwman, E. N. Kitova, M. Elaish, D. Kumawat, G. C. Daskhan, I. Tomris, L. Han, P. Chopra, T.-J. Yang, S. D. Willows, A. L. Mason, L. K. Mahal, T. L. Lowary, L. J. West, S.-T. D. Hsu, T. Hobman, S. M. Tompkins, G.-J. Boons, R. P. de Vries, M. S. Macauley and J. S. Klassen, Nat. Chem. Biol., 2022, 18, 81–90 CrossRef CAS PubMed
- V. Vantaku, S. R. Donepudi, C. R. Ambati, F. Jin, V. Putluri, K. Nguyen, K. Rajapakshe, C. Coarfa, V. L. Battula, Y. Lotan and N. Putluri, Oncotarget, 2017, 8, 95620–95631 CrossRef PubMed
- S. Groux-Degroote and P. Delannoy, Int. J. Mol. Sci., 2021, 22, 6145 CrossRef CAS PubMed
- C.-L. Schengrund, Int. J. Mol. Sci., 2020, 21, 5313 CrossRef CAS
- J. Wei, M. Hu, K. Huang, S. Lin and H. Du, Int. J. Mol. Sci., 2020, 21, 5983 CrossRef CAS
- E. Kakkis and D. Marsden, Mol. Genet. Metab., 2020, 130, 7–15 CrossRef CAS
- S. Bratulic, A. Limeta, S. Dabestani, H. Birgisson, G. Enblad, K. Stålberg, G. Hesselager, M. Häggman, M. Höglund, O. E. Simonson, P. Stålberg, H. Lindman, A. Bång-Rudenstam, M. Ekstrand, G. Kumar, I. Cavarretta, M. Alfano, F. Pellegrino, T. Mandel-Clausen, A. Salanti, F. Maccari, F. Galeotti, N. Volpi, M. Daugaard, M. Belting, S. Lundstam, U. Stierner, J. Nyman, B. Bergman, P.-H. Edqvist, M. Levin, A. Salonia, H. Kjölhede, E. Jonasch, J. Nielsen and F. Gatto, Proc. Natl. Acad. Sci. U. S. A., 2022, 119, e2115328119 CrossRef CAS PubMed
- P.-F. Zhang, Z.-Y. Wu, W.-B. Zhang, Y.-Q. He, K. Chen, T.-M. Wang, H. Li, H. Zheng, D.-H. Li, D.-W. Yang, T. Zhou, C.-M. Deng, Y. Liao, W.-Q. Xue, L.-J. Cao, X.-Z. Li, J.-B. Zhang, S.-Q. Dong, F. Wang, M.-Q. Zheng, W.-L. Zhang, J. Mu and W.-H. Jia, Nat. Commun., 2023, 14, 645 CrossRef CAS
- S. S. Pinho and C. A. Reis, Nat. Rev. Cancer, 2015, 15, 540–555 CrossRef CAS
- M. Yamashina, M. Akita, T. Hasegawa, S. Hayashi and M. Yoshizawa, Sci. Adv., 2017, 3, e1701126 CrossRef PubMed
- Y. Xiong, X. Li, M. Li, H. Qin, C. Chen, D. Wang, X. Wang, X. Zheng, Y. Liu, X. Liang and G. Qing, J. Am. Chem. Soc., 2020, 142, 7627–7637 CrossRef CAS
- S. Tommasone, F. Allabush, Y. K. Tagger, J. Norman, M. Köpf, J. H. R. Tucker and P. M. Mendes, Chem. Soc. Rev., 2019, 48, 5488–5505 RSC
- S. A. Byron, K. R. Van Keuren-Jensen, D. M. Engelthaler, J. D. Carpten and D. W. Craig, Nat. Rev. Genet., 2016, 17, 257–271 CrossRef CAS PubMed
- Y. Yoshida, J. I. Furukawa, S. Naito, K. Higashino, Y. Numata and Y. Shinohara, Proteomics, 2016, 16, 2747–2758 CrossRef CAS
- T. R. McKitrick, D. Eris, N. Mondal, R. P. Aryal, N. McCurley, J. Heimburg-Molinaro and R. D. Cummings, Biochemistry, 2020, 59, 3111–3122 CrossRef CAS
- E. Kaltgrad, S. Sen Gupta, S. Punna, C.-Y. Huang, A. Chang, C.-H. Wong, M. G. Finn and O. Blixt, ChemBioChem, 2007, 8, 1455–1462 CrossRef CAS
- E. Sterner, N. Flanagan and J. C. Gildersleeve, ACS Chem. Biol., 2016, 11, 1773–1783 CrossRef CAS PubMed
- H. Koprowski, Z. Steplewski, K. Mitchell, M. Herlyn, D. Herlyn and P. Fuhrer, Somatic Cell Genet., 1979, 5, 957–972 CrossRef CAS PubMed
- O. Haji-Ghassemi, R. J. Blackler, N. Martin Young and S. V. Evans, Glycobiology, 2015, 25, 920–952 CrossRef CAS
- J. C. Paulson, O. Blixt and B. E. Collins, Nat. Chem. Biol., 2006, 2, 238–248 CrossRef CAS PubMed
- K. Van Roey, B. Uyar, R. J. Weatheritt, H. Dinkel, M. Seiler, A. Budd, T. J. Gibson and N. E. Davey, Chem. Rev., 2014, 114, 6733–6778 CrossRef CAS PubMed
- A. Pincetic, S. Bournazos, D. J. DiLillo, J. Maamary, T. T. Wang, R. Dahan, B.-M. Fiebiger and J. V. Ravetch, Nat. Immunol., 2014, 15, 707–716 CrossRef CAS PubMed
- S. L. Sazinsky, R. G. Ott, N. W. Silver, B. Tidor, J. V. Ravetch and K. D. Wittrup, Proc. Natl. Acad. Sci. U. S. A., 2008, 105, 20167–20172 CrossRef CAS
- E. von Witting, S. Hober and S. Kanje, Bioconjugate Chem., 2021, 32, 1515–1524 CrossRef CAS PubMed
- K. Partyka, K. A. Maupin, R. E. Brand and B. B. Haab, Proteomics, 2012, 12, 2212–2220 CrossRef CAS
- R. Sawada, S.-M. Sun, X. Wu, F. Hong, G. Ragupathi, P. O. Livingston and W. W. Scholz, Clin. Cancer Res., 2011, 17, 1024–1032 CrossRef CAS PubMed
- M. S. Pereira, I. Alves, M. Vicente, A. Campar, M. C. Silva, N. A. Padrão, V. Pinto, Â. Fernandes, A. M. Dias and S. S. Pinho, Front. Immunol., 2018, 9, 2754 CrossRef PubMed
- L. G. Karacosta, J. C. Fisk, J. Jessee, S. Tati, B. Turner, D. Ghazal, R. Ludwig, H. Johnson, J. Adams, M. Sajjad, S. Koury, R. Roy, J. R. Olson and K. Rittenhouse-Olson, Transl. Oncol., 2018, 11, 450–466 CrossRef
- E. Sterner, N. Flanagan and J. C. Gildersleeve, ACS Chem. Biol., 2016, 11, 1773–1783 CrossRef CAS PubMed
- J. Hirabayashi, M. Yamada, A. Kuno and H. Tateno, Chem. Soc. Rev., 2013, 42, 4443–4458 RSC
- R. Mody, S. H. a Joshi and W. Chaney, J. Pharmacol. Toxicol. Methods, 1995, 33, 1–10 CrossRef CAS
- A. Pusztai and G. Grant, Methods Mol. Med., 1998, 9, 505–514 CAS
- W. I. Weis and K. Drickamer, Annu. Rev. Biochem., 1996, 65, 441–473 CrossRef CAS
- A. Dessen, D. Gupta, S. Sabesan, C. F. Brewer and J. C. Sacchettini, Biochemistry, 1995, 34, 4933–4942 CrossRef CAS
- S. R. Stowell, C. M. Arthur, P. Mehta, K. A. Slanina, O. Blixt, H. Leffler, D. F. Smith and R. D. Cummings, J. Biol. Chem., 2008, 283, 10109–10123 CrossRef CAS PubMed
- F. Teillet, B. Dublet, J. P. Andrieu, C. Gaboriaud, G. J. Arlaud and N. M. Thielens, J. Immunol., 2005, 174, 2870–2877 CrossRef CAS PubMed
- R. Arango, E. Rodriguez-Arango, R. Adar, D. Belenky, F. G. Loontiens, S. Rozenblatt and N. Sharon, FEBS Lett., 1993, 330, 133–136 CrossRef CAS PubMed
- N. Nishi, A. Itoh, A. Fujiyama, N. Yoshida, S.-I. Araya, M. Hirashima, H. Shoji and T. Nakamura, FEBS Lett., 2005, 579, 2058–2064 CrossRef CAS PubMed
- P. Adhikari, K. Bachhawat-Sikder, C. J. Thomas, R. Ravishankar, A. A. Jeyaprakash, V. Sharma, M. Vijayan and A. Surolia, J. Biol. Chem., 2001, 276, 40734–40739 CrossRef CAS
- N. Nishi, A. Itoh, A. Fujiyama, N. Yoshida, S.-I. Araya, M. Hirashima, H. Shoji and T. Nakamura, FEBS Lett., 2005, 579, 2058–2064 CrossRef CAS
- R. Liang, J. Loebach, N. Horan, M. Ge, C. Thompson, L. Yan and D. Kahne, Proc. Natl. Acad. Sci. U. S. A., 1997, 94, 10554–10559 CrossRef CAS
- A. Varki, Glycobiology, 2017, 27, 3–49 CrossRef CAS
- S. S. Komath, M. Kavitha and M. J. Swamy, Org. Biomol. Chem., 2006, 4, 973–988 RSC
- T. Hatakeyama, Methods Mol. Biol., 2020, 2132, 159–164 CrossRef CAS PubMed
- L. D. Powell and A. Varki, J. Biol. Chem., 1994, 269, 10628–10636 CrossRef CAS
- P. B. Bhanushali, S. B. Badgujar, M. M. Tripathi, S. Gupta, V. Murthy, M. V. Krishnasastry and C. P. Puri, Int. J. Biol. Macromol., 2016, 86, 468–480 CrossRef CAS PubMed
- F. Tobola, E. Sylvander, C. Gafko and B. Wiltschi, Interface Focus, 2019, 9, 20180072 CrossRef
- Z. Liu and H. He, Acc. Chem. Res., 2017, 50, 2185–2193 CrossRef CAS PubMed
- B. J. Graham, I. W. Windsor, B. Gold and R. T. Raines, Proc. Natl. Acad. Sci. U. S. A., 2021, 118, e2013691118 CrossRef CAS
- H. Xiao, W. Chen, J. M. Smeekens and R. Wu, Nat. Commun., 2018, 9, 1692 CrossRef
- H. Wang, Z. Bie, C. Lü and Z. Liu, Chem. Sci., 2013, 4, 4298–4303 RSC
- Y. Chen, S. Wang, J. Ye, D. Li, Z. Liu and X. Wu, Nanoscale, 2014, 6, 9563–9567 RSC
- P. Li, S. Xu, Y. Han, H. He and Z. Liu, Chem. Sci., 2023, 14, 2553–2561 RSC
- J. Zhou and J. Rossi, Nat. Rev. Drug Discovery, 2017, 16, 181–202 CrossRef CAS
- A. M. Yoshikawa, A. Rangel, T. Feagin, E. M. Chun, L. Wan, A. Li, L. Moeckl, D. Wu, M. Eisenstein, S. Pitteri and H. T. Soh, Nat. Commun., 2021, 12, 7106 CrossRef CAS PubMed
- A. V. Lakhin, V. Z. Tarantul and L. V. Gening, Acta Nat., 2013, 5, 34–43 CrossRef CAS
- G. Wulff and A. Sarhan, Angew. Chem., Int. Ed. Engl., 1972, 84, 364 CrossRef
- G. Wulff, Angew. Chem., Int. Ed. Engl., 1995, 34, 1812–1832 CrossRef CAS
- H. Shi, W. B. Tsai, M. D. Garrison, S. Ferrari and B. D. Ratner, Nature, 1999, 398, 593–597 CrossRef CAS PubMed
- M. Zangiabadi and Y. Zhao, J. Am. Chem. Soc., 2022, 144, 17110–17119 CrossRef CAS
- A. J. Kadhem, G. J. Gentile and M. M. Fidalgo de Cortalezzi, Molecules, 2021, 26, 6233 CrossRef CAS PubMed
- A. Jahanban-Esfahlan, L. Roufegarinejad, R. Jahanban-Esfahlan, M. Tabibiazar and R. Amarowicz, Talanta, 2020, 207, 120317 CrossRef CAS PubMed
- S. Akgönüllü and A. Denizli, J. Pharm. Biomed. Anal., 2023, 225, 115213 CrossRef
- X. Huang, W. Zhang, Z. Wu, H. Li, C. Yang, W. Ma, A. Hui, Q. Zeng, B. Xiong and Z. Xian, J. Mol. Model., 2020, 26, 198 CrossRef CAS PubMed
- K. Smolinska-Kempisty, A. Guerreiro, F. Canfarotta, C. Cáceres, M. J. Whitcombe and S. Piletsky, Sci. Rep., 2016, 6, 37638 CrossRef CAS PubMed
- R. Xing, S. Wang, Z. Bie, H. He and Z. Liu, Nat. Protoc., 2017, 12, 964–987 CrossRef CAS
- W. Li, Q. Zhang, Y. Wang, Y. Ma, Z. Guo and Z. Liu, Anal. Chem., 2019, 91, 4831–4837 CrossRef CAS
- A. Poma, H. Brahmbhatt, H. M. Pendergraff, J. K. Watts and N. W. Turner, Adv. Mater., 2015, 27, 750–758 CrossRef CAS PubMed
- X. Sun, Y. Jian, H. Wang, S. Ge, M. Yan and J. Yu, ACS Appl. Mater. Interfaces, 2019, 11, 16198–16206 CrossRef CAS PubMed
- X. Tu, P. Muhammad, J. Liu, Y. Ma, S. Wang, D. Yin and Z. Liu, Anal. Chem., 2016, 88, 12363–12370 CrossRef CAS
- S. Diken Gür, M. Bakhshpour and A. Denizli, New J. Chem., 2022, 46, 7263–7270 RSC
- T. Rebelo, R. Costa, A. Brandao, A. F. Silva, M. G. F. Sales and C. M. Pereira, Anal. Chim. Acta, 2019, 1082, 126–135 CrossRef CAS
- J. Li, X. Ma, M. Li and Y. Zhang, Biosens. Bioelectron., 2018, 99, 438–442 CrossRef CAS
- J. A. Ribeiro, C. M. Pereira, A. F. Silva and M. G. F. Sales, Biosens. Bioelectron., 2018, 109, 246–254 CrossRef CAS PubMed
- L. Zhou, Y. Wang, R. Xing, J. Chen, J. Liu, W. Li and Z. Liu, Biosens. Bioelectron., 2019, 145, 111729 CrossRef CAS PubMed
- K. Luo, C. Zhao, Y. Luo, C. Pan and J. Li, Biosens. Bioelectron., 2022, 216, 114672 CrossRef CAS PubMed
- J. Pang, P. Li, H. He, S. Xu and Z. Liu, Chem. Sci., 2022, 13, 4589–4597 RSC
- J. W. Hickey, E. K. Neumann, A. J. Radtke, J. M. Camarillo, R. T. Beuschel, A. Albanese, E. McDonough, J. Hatler, A. E. Wiblin, J. Fisher, J. Croteau, E. C. Small, A. Sood, R. M. Caprioli, R. M. Angelo, G. P. Nolan, K. Chung, S. M. Hewitt, R. N. Germain, J. M. Spraggins, E. Lundberg, M. P. Snyder, N. L. Kelleher and S. K. Saka, Nat. Methods, 2022, 19, 284–295 CrossRef CAS
- D. M. Krolewski, V. Kumar, B. Martin, R. Tomer, K. Deisseroth, R. M. Myers, A. F. Schatzberg, F. S. Lee, J. D. Barchas, W. E. Bunney, H. Akil and S. J. Watson, Jr., Brain Struct. Funct., 2018, 223, 987–999 CrossRef CAS
- Z. El-Schich, M. Abdullah, S. Shinde, N. Dizeyi, A. Rosén, B. Sellergren and A. G. Wingren, Tumour Biol., 2016, 37, 13763–13768 CrossRef CAS
- M. Kimani, S. Beyer, Z. El-Schich, K. Gawlitza, A. Gjörloff-Wingren and K. Rurack, ACS Appl. Polym. Mater., 2021, 3, 2363–2373 CrossRef CAS
- S. Shinde, Z. El-Schich, A. Malakpour, W. Wan, N. Dizeyi, R. Mohammadi, K. Rurack, A. Gjörloff Wingren and B. Sellergren, J. Am. Chem. Soc., 2015, 137, 13908–13912 CrossRef CAS PubMed
- W. Zhang, J. Kang, P. Li, L. Liu, H. Wang and B. Tang, Chem. Commun., 2016, 52, 13991–13994 RSC
- B. Demir, M. M. Lemberger, M. Panagiotopoulou, P. X. Medina Rangel, S. Timur, T. Hirsch, B. Tse Sum Bui, J. Wegener and K. Haupt, ACS Appl. Mater. Interfaces, 2018, 10, 3305–3313 CrossRef CAS PubMed
- M. Mahani, F. Mahmoudi, J. Fassihi, Z. Hasani and F. Divsar, Microchim. Acta, 2021, 188, 224 CrossRef CAS
- C. Dobie and D. Skropeta, Br. J. Cancer, 2021, 124, 76–90 CrossRef CAS PubMed
- S. Wang, Y. Wen, Y. Wang, Y. Ma and Z. Liu, Anal. Chem., 2017, 89, 5646–5652 CrossRef CAS PubMed
- S. Wang, D. Yin, W. Wang, X. Shen, J.-J. Zhu, H.-Y. Chen and Z. Liu, Sci. Rep., 2016, 6, 22757 CrossRef CAS PubMed
- X. H. Ren, H. Y. Wang, S. Li, X. W. He, W. Y. Li and Y. K. Zhang, Talanta, 2021, 223, 121706 CrossRef CAS
- S. Jiang, T. Wang, S. Behren, U. Westerlind, K. Gawlitza, J. L. Persson and K. Rurack, ACS Appl. Nano Mater., 2022, 5, 17592–17605 CrossRef CAS
- Q. Zhu, Z. Shen, F. Chiodo, S. Nicolardi, A. Molinaro, A. Silipo and B. Yu, Nat. Commun., 2020, 11, 4142 CrossRef CAS
- B.-H. Li and X.-S. Ye, Curr. Opin. Chem. Biol., 2020, 58, 20–27 CrossRef CAS PubMed
- S. Wang, Y. Yang, Q. Zhu, G.-Q. Lin and B. Yu, Curr. Opin. Chem. Biol., 2022, 69, 102154 CrossRef CAS
- K. Madhusudan Makwana and R. Mahalakshmi, Protein Sci., 2015, 24, 1920–1933 CrossRef CAS
- F. A. Quiocho, Annu. Rev. Biochem., 1986, 55, 287–315 CrossRef CAS
- M. Wimmerová, S. Kozmon, I. Nečasová, S. K. Mishra, J. Komárek and J. Koča, PLoS One, 2012, 7, e46032 CrossRef
- A. Varki, Nature, 2013, 497, 565 CrossRef CAS PubMed
- L. Zhu, Y. Xu, S. Kang, B. Lin, C. Zhang, Z. You, H. Lin, C. Yang and Y. Song, Small Methods, 2022, 6, e2200549 CrossRef
- T. Suzuki, Nat. Rev. Mol. Cell Biol., 2021, 22, 375–392 CrossRef CAS
- W. Tyagi, V. Pandey and Y. R. Pokharel, Cancer Gene Ther., 2023, 30, 641–646 CrossRef CAS
- C. A. Borrebaeck, Nat. Rev. Cancer, 2017, 17, 199–204 CrossRef CAS
- B. H. Park, Y. T. Kim, J. H. Jung and T. S. Seo, Microchim. Acta, 2014, 181, 1655–1668 CrossRef CAS
- C. S. Kosack, A. L. Page and P. R. Klatser, Bull. W. H. O., 2017, 95, 639–645 CrossRef PubMed
-
E. Orrantia-Borunda, P. Anchondo-Nuñez, L. E. Acuña-Aguilar, F. O. Gómez-Valles and C. A. Ramírez-Valdespino, in Breast Cancer, ed. H. N. Mayrovitz, Exon Publications, 2022 Search PubMed
- J. Pan, Y. Hu, S. Sun, L. Chen, M. Schnaubelt, D. Clark, M. Ao, Z. Zhang, D. Chan, J. Qian and H. Zhang, Nat. Commun., 2020, 11, 6139 CrossRef CAS
- M. Tonutti, G. Gras and G. Z. Yang, Artif. Intell. Med., 2017, 80, 39–47 CrossRef PubMed
- L. M. Petrick and N. Shomron, Cell Rep. Phys. Sci., 2022, 3, 100978 CrossRef CAS
| This journal is © The Royal Society of Chemistry 2024 |